Pour se former tout au long de la vie, il faut être capable d’apprendre et pour cela de mobiliser son attention. La recherche en neurosciences suggère que ces deux fonctions du cerveau sont certes très performantes par leur capacité mais présentent également des limites structurelles. Nos travaux en étudient certaines modulations, notamment le rôle de la dominance et de la soumission ou encore l’interaction avec les modes mentaux.
Intégrer dans les modalités pédagogiques ces réalités de base pour mieux les gérer, valoriser des leviers d’action innovants, pourrait avoir un impact notable sur les résultats obtenus à tous les stades de l’apprentissage humain : réduction de l’échec, du stress de l’apprenant… comme de celui de l’enseignant, aussi bien dans l’éducation générale, les formations universitaires et professionnelles initiales, que dans la formation continue proposée aux entreprises et aux particuliers…
 Le programme NeuroTEACH consiste en une série d’études en neurosciences cognitives explorant le rôle modulateur des relations sociales, notamment celles de dominance et de soumission, sur les mécanismes de l’attention et l’apprentissage.
Le programme NeuroTEACH consiste en une série d’études en neurosciences cognitives explorant le rôle modulateur des relations sociales, notamment celles de dominance et de soumission, sur les mécanismes de l’attention et l’apprentissage.
Il résulte de la fructueuse collaboration entre 2010 et 2017 (cf. § Liste de publications) entre l’IME (E. Monfardini, J. Fradin) et le Centre de Recherche en Neurosciences de Lyon, un des grands pôles neuroscientifiques de la recherche publique en France (Martine Meunier, équipe ImpAct, Inserm – CNRS – Université Lyon 1).
L’objectif principal du projet était d’explorer des moyens d’optimiser l’attention et l’apprentissage et donc le potentiel cognitif des individus sans recours aux médicaments.
Les objectifs secondaires étaient les transferts de connaissance sur les mécanismes neurobiologiques de l’apprentissage et de l’attention issues des études NeuroTEACH pour améliorer les pratiques éducatives en contexte scolaire mais aussi toute la vie, notamment en contexte professionnel, afin de répondre aux besoins actuels des individus et des organisations d’optimisation de leurs performances et de leur compétitivité.
Quelques communiqués de presse à lire :
- Actualités INSERM « Le mimétisme nécessaire à l’apprentissage » 14 avril 2014, communiqué repris par de nombreux autres sites Web médias http://www.inserm.fr/actualites/rubriques/actualites-recherche/le-mimetisme-necessaire-a-l-apprentissage
- Institut de NeuroCognitisme : « Neuropédagogie: Pourquoi les sciences du cerveau changent la donne en formation? » 10 janvier 2013http://www.neurocognitivisme.fr/fr/actualite.php?doc_id=641
- Science News : « Being watched can boost productivity » 17 Avril 2015 https://www.sciencenews.org/article/being-watched-can-boost-productivity?mode=topic&context=69&tgt=nr
Mise à jour : mai 2017
Monfardini Elisabetta, Projets ISCI: contexte théorique et état de l’art
Contexte théorique
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant des spécialistes dans les domaines des neurosciences cognitives, de la psychologie sociale et de l’éthologie comparée. Plus particulièrement, les travaux « ISCI » portent sur l’influence du contexte et des interactions sociales sur le fonctionnement cognitif et cérébral des individus, et ils utilisent les méthodes scientifiques des neurosciences et de la psychologie sociale (Necka et al., 2015; Adolphs, 2010; voir aussi Ochsner & Lieberman, 2001).
Ces programmes de recherche fondamentale s’inscrivent dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres. Dans l’approche proposée, le PG aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études décrites ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est extrêmement stable car présent chez différentes espèces et tout au long du développement.
Le but du présent axe de recherche est d’explorer la modulation sociale de la cognition, ses substrats neuronaux, les mécanismes neurocognitifs qui la gouvernent, ainsi que son évolution au cours de l’ontogénèse et de la phylogénèse. Nous explorerons l’influence du contexte social sur nos comportements et habilités afin de tester nos hypothèses autour de la cognition sociale et du fonctionnement cérébral qui en est responsable. Nous visons tout particulièrement à répondre aux questions suivantes :
- Sommes-nous en quelque sorte « programmés » pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (cf. Fig.1, volet Fondamental) ;
- Quelles modulations sociales sont des tendances stables et anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes? (cf. Fig.1, volet Fondamental) ;
- Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale? (cf. Fig.1, volet Bases Neurales) ;
- Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions ont été abordées via plusieurs approches complémentaires :
- Une approche comportementale comparée qui met en relation (i) le comportement social de l’homme à celui de l’animal, dans le but d’étudier la phylogenèse de l’influence sociale et d’isoler ses composantes fondamentales ; (ii) le comportement de l’homme adulte à celui de l’enfant, afin de suivre le développement de l’influence sociale et ses répercussions notamment sur l’éducation et l’acquisition de connaissances;
- Deux techniques d’imagerie cérébrale: la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) (toutes deux permettent une visualisation non invasive de l’activité du cerveau) appliquées à l’homme et au singe macaque;
- L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc.
D’un point de vue applicatif, les résultats des recherches ISCI iront, d’une part, étoffer les contenus de l’approche Neurocognitive proposée par l’IME et, d’autre part, pourront faire l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, étudiants, etc.) fait l’objet d’un projet communément mené par le Centre de Recherche en Neurosciences de Lyon (INSERM U1028, CNRS UMR5292) et l’IME, dénommé NeuroTeach. Ce projet est désormais promu par la Fondation Neurodis, reconnue d’utilité publique (cf. http://www.fondation-neurodis.org/newsletters/novembre2015/). En définitive, reposant sur une solide collaboration scientifique entre l’IME et les institutions de recherche publique comme le CNRL, les projets ISCI ont l’ambition de combiner les progrès des neurosciences fondamentales et les exigences commerciales d’entreprise, afin de proposer des applications dans le domaine notamment de la pédagogie.
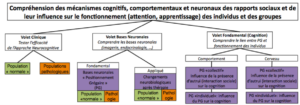
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est la position/rang social/e (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres.
État de l’art
La psychologie a fourni des données expérimentales sur la modulation sociale du fonctionnement cognitif individuel. Les études en psychologie sociale témoignent, par exemple, du fait que la présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives. Il est de tradition depuis Allport (1924) de parler de « facilitation sociale », pour désigner ce phénomène, bien qu’il ait été montré par la suite (Zajonc, 1965; Bond & Titus, 1983; Guerin & Innes, 2009) que la présence des autres peut avoir des conséquences soit positives (pour des réponses déjà apprises), soit négatives (pour des réponses nouvelles). Ce phénomène de facilitation/inhibition sociale de la cognition concerne une large palette de capacités: des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements, cf. Bond & Titus, 1983), aux plus complexes comme l’attention (e.g. dans la tâche de Stroop, Huguet et al., 2009), la prise de décision (e.g. Asch, 1956), ou l’apprentissage (trouver le bon mouvement associé à un stimulus est plus difficile lorsque nous sommes observés, Belletier et al., 2015; Bounoua et al., 2009). . La psychologie comparée a montré, quant à elle, la vaste représentation phylogénétique de l’influence des autres. Les phénomène de facilitation/inhibition sociale, tout comme le phénomène de « social buffering » (i.e. la meilleure récupération suite à des expériences aversives des sujets lorsqu’ils vivent ensemble; Kikusui et al., 2006) se retrouvent chez nombre d’espèces animales qu’il s’agisse d’insectes, de rongeurs, ou de primates non-humains et humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire (nous mangeons plus en groupe que seuls, cf. Herman 2015). Dans le règne animal, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Des situations sociales relativement simples, donc, peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales et cognitives. La modulation sociale de la cognition trouve son ancrage dans l’activité cérébrale. Des études en neuroimagerie chez l’homme ont mis en évidence, en effet, que l’activité du cerveau liée à des fonctions supérieures telles que la prise de décision (Rilling & Sanfey, 2011) ou l’apprentissage (Monfardini et al., 2013) est modulée par le contexte social. L’électrophysiologie chez le singe macaque (le modèle animal par excellence des neurosciences sociales) suggère que l’information sociale modifie l’activité électrique des neurones en plusieurs aires corticales (Azzi et al., 2012; Rizzolatti & Fogassi, 2014). Dans une série d’études, Fujii, Hihara et Iriki (2007, 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone. Par la suite, des études de neuroimagerie chez l’homme ont mis en évidence un changement de l’activité cérébrale selon que les participants jouaient avec une autre personne, par rapport à un ordinateur ou seuls (Nawa et al., 2008;. Fareri et al., 2012; Kätsyri et al., 2013).
La dimension sociale semble donc faire donc partie intégrante des mécanismes cérébraux de base. Pourtant (1) les mécanismes cérébraux qui modulent nos capacités en présence d’autrui demeurent inconnus, et (2) le système éducatif (école, formation continue) ne prend pas toujours cela en considération quand il choisit ses outils pédagogiques, et ceci vaut aussi bien en contexte scolaire que dans le monde professionnel. Certaines pratiques récurrentes à l’école sont susceptibles de détériorer les performances des élèves. Nous avons démontré que les adultes (hommes ou singes qu’ils soient) apprennent aisément des erreurs d’autrui, alors que l’apprentissage par les succès des autres n’est pas donné à tout le monde et dépend de certaines caractéristiques individuelles (Monfardini et al., 2012). Dans certaines conditions, apprendre de quelqu’un de trop performant peut même être délétère par rapport à l’apprentissage individuel (Belletier et al., 2015; Bounoua et al., 2009), et les caractéristiques du modèle exercent une influence drastique sur l’apprentissage (Monfardini et al, 2014). Si répliqués chez l’enfant, ces résultats impliquent que choisir le meilleur élève (qui ne fait donc pas d’erreurs) comme modèle pour les autres peut ne pas être la meilleure stratégie pour la transmission de connaissances dans une classe. Les projets ISCI partent du constat que connaître l’évolution (onto- et phylogénétique) de l’influence des autres sur nos capacités cognitives peut donc ouvrir de prometteuses perspectives dans le domaine de la pédagogie. De récentes avancées en neuroscience ont déjà fourni d’importantes suggestions pour l’éducation. La neuropédagogie a proposé, par exemple, de outils pratiques de «contrôle cognitif » issus des connaissances neuroscientifiques pour aider les enfants à inhiber leur tendance à donner des réponses « automatiques » (biais d’automatisme; Diamond & Lee, 2011).
En définitive, la littérature existante en neurosciences et psychologie témoigne du fait que notre cerveau tient en compte des autres pour moduler sa propre activité. En revanche, elle ne permet pas de répondre aux questions suivantes : pourquoi et dans quelle conditions, nos comportements et nos habilités sont modifiées à cause de la présence d’autrui? Comment varie cette influence sociale au cours du développement, et quelle est son origine phylogénétique? Et encore, par quels mécanismes neurobiologiques passe cette influence sociale? Quelles sont les dynamiques cérébrales qui sont influencées par les autres, et qui influencent nos relations avec les autres? Comment intégrer ces connaissances afin d’améliorer l’éducation et donc le potentiel d’habilités cognitives des personnes? A travers de la série d’expériences ISCI, en cours ou menées avec succès au cours des 6 dernières années, nous tâchons de répondre à toutes ces questions comportementales (cf. par exemple ISCI3, ISCI9, ISCI10, ISCI12 et ISCI13), et nous laissons notamment aux projets ISCI4 (complété), ISCI7 (complété), ISCI11 et ISCI13 la tâche d’explorer les mécanismes neurobiologiques de l’influence sociale.
Articles scientifiques ISCI parus en 2016:
- Monfardini E, Reynaud AJ, Prado J, Meunier M (2016). Social modulation of cognition: lessons from rhesus macaques relevant to education. Neurosci. Biobehav. Rev. IF 10.1
- Guedj C, Monfardini E, Reynaud AJ, Farnè A, Meunier M, Hadj-Bouziane F (2016). Boosting norepinephrine transmission triggers flexible reconfiguration of brain networks at rest. Cereb Cortex. doi: 10.1093/cercor/bhw262. IF 8.3
- Wicker B, Monfardini E, Royet JP (2016). Olfactory processing in adults with Autism Spectrum Disorders. Mol Autism. 19;7:4. doi: 10.1186/s13229-016-0070-3. IF 4.96
Articles scientifiques ISCI soumis en 2016:
- Guedj, C., Reynaud, A., Monfardini, E., Salemme, R., Farne, A., Meunier, M., & Hadj-Bouziane, F. (under revision) Boosting norepinephrine transmission optimizes response strategy. Journal of Neuroscience.
Bibliographie
Adolphs, R. (2010). Conceptual challenges and directions for social neuroscience. Neuron, 65, 752-67.
Allport, F. H. (1924) Social psychology. Boston: Houghton Mifflin.
Asch, S. E. (1956). Studies of independence and conformity: I. A minority of one against a unanimous majority. Psychological monographs: General and applied, 70, 1.
Azzi, J. C., Sirigu, A., & Duhamel, J. R. (2012). Modulation of value representation by social context in the primate orbitofrontal cortex. Proceedings of the National Academy of Sciences, 109(6), 2126-2131.
Belletier, C., Davranche, K., Tellier, I. S., Dumas, F., Vidal, F., Hasbroucq, T., & Huguet, P. (2015). Choking under monitoring pressure: being watched by the experimenter reduces executive attention. Psychonomic bulletin & review, 22(5), 1410-1416.
Bond Jr, C. F., & Titus, L. J. (1983). Social Facilitation: A Meta-Analysis of 241 Studies. Psychological Bulletin, 94, 265-292.
Bounoua L, Monfardini E, Meunier M, Wicker B, Brovelli A, Boussaoud D, Cury F, Huguet P (2009). Learning by Observation: The role of social comparison and related ability inferences. ESCOP, Poland.
Diamond, A., & Lee, K. (2011). Interventions shown to aid executive function development in children 4 to 12 years old. Science, 333(6045), 959-964.
Dindo, M., Whiten, A., & De Waal, F. (2009). Social facilitation of exploratory foraging behavior in capuchin monkeys (Cebus apella). American Journal of Primatology, 71(5), 419-426.
Fareri, D. S., Niznikiewicz, M. A., Lee, V. K., & Delgado, M. R. (2012). Social network modulation of reward-related signals. The Journal of neuroscience, 32, 9045-9052.
Fujii, N., Hihara, S., & Iriki, A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PloS one, 2(4), e397.
Fujii, N., Hihara, S., & Iriki, A. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience, 3(3-4), 250-260.
Galloway, A. T., Addessi, E., Fragaszy, D. M., & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchins (Cebus apella): does it involve behavioral coordination?. International Journal of Primatology, 26(1), 181-189.
Guerin, B., & Innes, J. (2009). Social Facilitation. Cambridge: Cambridge University Press.
Huguet, P., Galvaing, M. P., Monteil, J. M., & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of personality and social psychology, 77, 1011.
Kätsyri, J., Hari, R., Ravaja, N., & Nummenmaa, L. (2012). The opponent matters: elevated fMRI reward responses to winning against a human versus a computer opponent during interactive video game playing. Cerebral Cortex, bhs259.
Kikusui, T., Winslow, J. T., & Mori, Y. (2006). Social buffering: relief from stress and anxiety. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 361(1476), 2215-2228.
Monfardini, E., Gaveau, V., Boussaoud, D., Hadj-Bouziane, F., & Meunier, M. (2012). Social Learning as a Way to Overcome Choice-Induced Preferences? Insights from Humans and Rhesus Macaques. Frontiers in Neuroscience, 6.
Monfardini, E., Gazzola, V., Boussaoud, D., Brovelli, A., Keysers, C., & Wicker, B. (2013). Vicarious neural processing of outcomes during observational learning. PloS one, 8(9), e73879.
Monfardini, E., Hadj-Bouziane, F., & Meunier, M. (2014). Model-observer similarity, error modeling and social learning in rhesus macaques. PloS one, 9(2), e89825.
Nawa, N. E., Nelson, E. E., Pine, D. S., & Ernst, M. (2008). Do you make a difference? Social context in a betting task. Social cognitive and affective neuroscience, 3, 367-376.
Necka, E. A., Cacioppo, S., & Cacioppo, J. T. (2015). Social Neuroscience of the Twenty-First Century. In J. D. Wright (Ed.) International Encyclopedia of the Social & Behavioral Sciences (2nd edition), (Volume 22, p.485-488). http://dx.doi.org/10.1016/B978-0-08-097086-8.56020-6
Ochsner, K. N., & Lieberman, M. D. (2001). The emergence of social cognitive neuroscience. American Psychologist, 56, 717.
Reynaud, A. J., Guedj, C., Hadj-Bouziane, F., Meunier, M., & Monfardini, E. (2015). Social facilitation of cognition in rhesus monkeys: audience vs. coaction. Frontiers in behavioral neuroscience, 9.
Rilling, J. K., & Sanfey, A. G. (2011). The neuroscience of social decision-making. Annual review of psychology, 62, 23-48.
Rizzolatti, G., & Fogassi, L. (2014). The mirror mechanism: recent findings and perspectives. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1644).
Visalberghi, E., & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour, 60(1), 69-76.
Visalberghi, E., & Addessi, E. (2001). Acceptance of novel foods in capuchin monkeys: do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour, 62(3), 567-576.
Visalberghi, E., & Fragaszy, D. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour, 49(4), 1089-1095.
Zajonc, R. B. (1965). Social facilitation. Science. 149: 269-274
Mise à jour : mai 2016
| NOM DU PROJET : ISCI2 -Principe de similarité et apprentissage par observation : étude comportementale chez le singe macaque rhésus (ISCI2). |
| Date de début : 2011 | Date de fin : 2014 |
| R. Psychologie – Neurosciences |
1. Objectifs du projet
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant les neurosciences cognitives, la psychologie sociale, l’éthologie comparée et la neuroendocrinologie. Plus particulièrement, ces travaux portent sur l’influence des interactions sociales sur le fonctionnement cognitif et cérébral individuel. Cela à travers les méthodes scientifiques des neurosciences fondamentales et de la psychologie (Ochsner & Lieberman, 2001).
Le présent axe de recherche s’inscrit dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et d’intégration/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établit avec les autres. Dans l’approche proposée, le PG aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études exposées ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est phylogénétiquement ancien. Étudier cette influence nous permettra donc d’explorer nos hypothèses en termes de cognition sociale et du fonctionnement cérébral qui en est responsable. D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Le but du présent projet de recherche est d’explorer la modulation sociale de la cognition, les mécanismes qui la gouvernent, son évolution au cours de la phylogénèse, ainsi que ses substrats neuronaux. Il vise en particulier à répondre aux questions suivantes :
– Sommes-nous en quelque sorte ‘programmés’ pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (cf. Fig.1, volet Fondamental) ;
– Quelles modulations sociales sont des tendances phylogénétiquement anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes ? (cf. Fig.1, volet Fondamental) ;
– Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale ? (cf. Fig.1, volet Bases Neurales) ;
– Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions sont abordées via trois approches complémentaires :
– Une approche comportementale comparée confrontant le comportement de l’homme à celui de l’animal mis dans une situation similaire ; (cf. Projets ISCI2 et ISCI3)
– Deux techniques d’imagerie cérébrale humaine : la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) appliquées au macaque et à l’homme (toutes deux permettent une visualisation non invasive de l’activité du cerveau) ; cf Projets ISCI4, ISCI6 et ISCI8.
– L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc. (cf. ISCI7).
D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
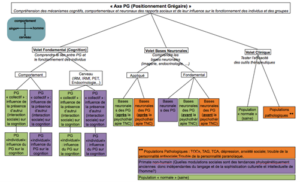
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est simplement la position sociale (« social ranking », en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres (« social comparison »).
2. État de l’art
La psychologie sociale a fourni des données expérimentales sur l’influence de la socialité sur le fonctionnement cognitif individuel. Ces études témoignent du fait que la simple présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives, consistant soit à les améliorer, soit à les détériorer (pour une revue récente de la littérature, voir Guerin & Innes, 2009). Le phénomène de modulation sociale concerne une large palette de capacités cognitives : des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements), aux plus complexes comme les capacités attentionnelles (e.g. dans la tâche de Stroop, Huguet et al., 1999), la prise de décision (Asch, 1956 ; Martin, 1998 ; ), ou l’apprentissage social de règles (Bounoua et al., 2009). En résumé, des situations sociales relativement simples peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales ou cognitives.
Les effets d’influence sociale ne sont pas une prérogative humaine. Des phénomènes de facilitation sociale ont aussi été démontrés chez les primates non-humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire. Par exemple, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Au niveau cérébral, une seule série d’études a jusqu’à présent donné quelques éléments pour répondre à cette question. Les auteurs (Fujii et al., 2007 ; 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone.
Comme nous l’avons précédemment dit, l’apprentissage social est également une composante fondamentale de la cognition humaine. Apprendre par observation d’autrui est vital pour l’homme (Castro & Toro, 2004). Sans cet apprentissage, chaque enfant devrait réapprendre chaque comportement laborieusement par lui-même et aucune transmission de savoir-faire ne serait possible. Selon Bandura (1965), ce type d’apprentissage est caractérisé par quatre étapes: l’attention, le maintien en mémoire, la reproduction du geste, et la motivation. La première étape, l’attention, est un processus crucial et peut être fortement influencé par les caractéristiques du modèle. Il a été démontré que des caractéristiques telles que l’attractivité, la similitude et la compétence perçue améliorent l’efficacité de l’apprentissage par observation (Brewer & Wann, 1998). En particulier, la similitude entre acteur et observateur serait la clé d’une transmission sociale des connaissances efficace. Il a été démontré, par exemple, que les enfants bénéficient plus de l’observation d’un pair novice (à qui ils peuvent s’identifier) que de la parfaite maîtrise d’un expert (donc perçu comme « trop supérieur » ). Chez les adolescents qui apprennent à écrire des textes, par exemple, les mauvais élèves apprennent mieux quand ils copient des modèles médiocres, tandis que les bons élèves apprennent mieux des modèles plus performants (Braaksma et al., 2002). De façon similaire, les adultes savent bénéficier de l’observation d’individus dont les performances sont légèrement supérieures quand ils s’agit d’apprendre des associations stimulus-récompense, mais au contraire ne bénéficient pas de l’observation d’individus perçus comme trop experts (Bounoua et al., 2009).
Des récentes découvertes de neurosciences corroborent ces données, en montrant une modulation des mécanismes cérébraux responsables de l’apprentissage social selon le dégrée de similarité et/ou affinité perçu entre modèle et apprenti (Mobbs et al., 2009 ; Leng & Zhou, 2010). Le principe de similarité, selon lequel plus les observateurs ressemblent aux modèles, meilleure est l’apprentissage social, semble valoir aussi pour les animaux non-humains, les singes (Bonnie et al., 2006) et les oiseaux (Schwab et al., 2008). Quand il s’agit de copier leur mère, les chimpanzés (Lonsdorf et al., 2004) et les capucins filles (Perry, 2009) sont meilleures de ce qui ne le sont les fils (garçons). Malgré leur intérêt, ces premières études d’éthologie comportementale et de psychologie comparative n’ont malheureusement aucune suite.
Le présent projet ISCI2 se base sur ce concept de similarité, bien connu en psychologie sociale et désormais assumé par les neurosciences cognitives, mais complètement ignoré dans le domaine de l’éthologie et des sciences de l’éducation. La question principale sous-jacente à ce projet a donc été de savoir si le principe de similarité a des origines phylogénétiquemment anciennes, ou s’il repose sur un biais cognitif qui est l’apanage de la culture humaine. Cette approche comparative a un clair avantage théorique mais elle apporte aussi des éléments pratiques dans les domaines de l’éducation et de la formation, aussi bien initiale que continue. En effet, comprendre les mécanismes de fonctionnement de l’apprentissage et de la transmission de connaissances, ses médiateurs et ses modérateurs qui transcendent toute spécificité d’espèce, permet de moduler les instruments éducatifs selon les caractéristiques intrinsèques de l’homme.
3. Aléas, incertitudes scientifiques, verrous technologiques
L’étude neuroscientifique de la modulation sociale de la cognition et, de façon plus générale, de la sphère sociale de notre existence, présente un grand intérêt mais aussi des défis techniques considérables.
Le défi essentiel de ce domaine est le contrôle d’un grand nombre de variables qui interviennent dans nos rapports sociaux et qui influencent notre « positionnement grégaire ». La complexité de ce domaine de recherche implique la multiplication des variables expérimentales à inclure dans nos protocoles expérimentaux et, par conséquent, des coûts considérables en termes de temps et de prix d’examens (notamment pour ce qui concerne les études neuroscientifiques d’imagerie). Cependant, les collaborations scientifiques sur lesquelles ces études reposent nous permettent de faire face aux problèmes liés à l’originalité de ce domaine d’étude. Ainsi, les résultats de ces études ont déjà fait l’objet de plusieurs publications dans des journaux scientifiques internationaux (Plos One, NeuroImage, Frontiers In Neurosciences, etc) et positionnent le Pôle de Recherche de l’IME dans le panorama scientifique international aussi bien dans le domaine de la Psychologie Cognitive que des Neurosciences.
L’étude ISCI2 implique l’utilisation d’un modèle animal, le primate non-humain. Ceci rentre dans le cadre d’une approche comparative finalisée à isoler les composantes fondamentales des comportements humains, qui s’expriment indépendamment de la culture et du langage. En particulier, le choix du modèle primate est motivé par l’intérêt que nous portons aux hiérarchies sociales et au positionnement grégaire. Or, les singes macaques passent la majeure partie de leur vie éveillée en présence de leurs congénères, et les rapports qu’ils instaurent au sein d’un groupe sont gérés par de fortes hiérarchies sociales. L’utilisation d’un modèle animal comporte des coûts et des aléas tout à fait différents de ceux auxquels on fait face pour les études chez l’homme. Tout d’abord, l’hébergement des animaux requiert du personnel et des infrastructures très coûteuses. Ensuite, la réalisation de protocoles expérimentaux impliquant des tâches cognitives requiert l’entrainement des animaux, qui se fait de façon progressive souvent sur des longues périodes (1 à 6 mois). Cet entrainement, qui comporte des travers les méthodes de conditionnement opérant bien connues en éthologie et psychologie comparative, demande des compétences particulières de la part du personnel impliqué dans l’étude. Le projet ISCI2 a été mené avec succès en collaboration avec le Centre de Recherche en Neurosciences de Lyon, qui possède une animalerie primate complètement rénovée et pouvant héberger un nombre important d’animaux. Le personnel INSERM qui y travaille a assuré le soin des animaux et leur hébergement conformément aux directives européennes pendant tout le déroulement du projet.
4. Travaux R&D réalisés, démarche expérimentale
Pour cette étude, nous avons adapté une tâche de résolution de problèmes issue d’une étude précédente (Meunier et al., 2007) pour répondre à cette question. Brièvement, sept macaques rhésus étaient testés lors qu’ils apprenaient de trois modèles différents: un congénère ‘novice’ de la tâche (i.e. qui à son tour était en train d’apprendre ; condition M = monkey) ; un humain ‘novice’ de la tâche (qui donc exécutait la tâche de la même façon que dans la condition M, y compris manger la récompense ; condition M-H = monkey-like) ; un humain qui montrait seulement la bonne ou la mauvaise réponse pour résoudre le problème, mais qui n’obtenait pas ni ne mangeait la récompense à chaque essai correct (condition EH = expert human).
Les résultats indiquent que le modèles M et M-L sont également efficaces pour apprendre une tâche associative stimulus-récompense. En revanche, l’homme expert qui ne mange pas la récompense (condition EH) se révèle inutile voire délétère par rapport à l’apprentissage individuel (cf. Fig. 2). Cela corrobore nos hypothèses, en laissant penser qu’un modèle d’apprentissage novice est plus bénéfique par rapport à un modèle expert et auquel il est difficile de s’identifier. S’appliquant aussi bien à l’homme qu’aux primates non-humains, le concept de similarité ne serait donc pas une caractéristique spécifique à l’homme mais un biais phylogénétiquement ancien qui témoigne, encore une fois, de l’effet que les dynamiques sociales peuvent avoir sur la cognition individuelle.
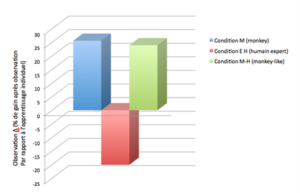
Figure 2. Résultats de l’étude 2. Un modèle d’apprentissage novice est plus bénéfique par rapport à un modèle expert et auquel il est difficile de s’identifier.
5. Indicateurs de R&D
L’article décrivant les résultats de cette étude en anglais a été publié dans le Journal ‘PlosOne’ en 2014 (Monfardini, Hadj-Bouziane & Meunier, 2014). Ces résultats ont également été présentés en conférence (Monfardini, 2014).
6. Acquisition de connaissances
Cette étude ouvre la voie à des réflexions théoriques et à des travaux empiriques dans le cadre de pédagogie et les sciences de l’éducation. Elle démontre en effet que, pour apprendre d’autrui, il faut se reconnaître dans l’autre et lui ressembler. Ceci pourrait remettre en cause certaines pratiques éducatives récurrentes, aussi bien en contexte scolaire que en formation continue : choisir le meilleur élève (qui ne fait donc pas d’erreurs et auquel il est plus difficile de s’identifier, comme modèle pour les autres peut ne pas être la meilleure stratégie pour la transmission de connaissances (Monfardini et al., 2012 ; http://www.inserm.fr/actualites/rubriques/actualites-recherche/le-mimetisme-necessaire-a-l-apprentissage). D’un point de vue applicatif, donc, les résultats de cette recherche seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach, et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
7. Ressources humaines
| Nom | Fonction dans le projet | Tâches relatives au projet | Nb total d’heures/jours affectées au projet |
| Elisabetta Monfardini | Chef de projet | Traitement des résultats et écriture de l’article (2013) | 346 heures |
8. Références bibliographiques
Asch, S.E. (1956). Studies of independence and conformity: I. A minority of ne against a unanimous majority. Psychological Monographs 70(9, Whole No. 416).
Bandura, A. (1965). Influence of models’ reinforcement contingencies on the acquisition of imitative responses. Journal of Personnality and Social Psychology, 1, 589-595.
Bonnie, K.E. & de Waal, F.B.M. (2006). Affiliation promotes transmission of a social custom: handclasp grooming among captive chimpanzees. Primates 47:27–34.
Bounoua, L., Cury, F., Monfardini, E., Meunier, M., Boussaoud, D., Brovelli, A. & Huguet, P. (2009). Learning by observation: The role of social comparison and related ability inferences. 16th Annual ESCOP conference, Jagiellonian University, Krakow.
Braaksma, M.H., Rijlaarsdam, G. & van den Bergh, H. (2002). Observational learning and the effects of model-observer similarity. Journal of Educational Psychology, 94(2), 405–415.
Brewer, K.R., Wann, D. L. (1998). Observational learning effectiveness as a function of model characteristics: Investigating the importance of social power. Social Behavior and Personality, 26(1), 1-10.
Castro, L. & Toro, M. (2004). The evolution of culture: From primate social learning to human culture. Proceedings of the National Academy of Sciences, 101: 10235-10240.
Dindo, M., Whiten, A. & de Waal, F. B. M. (2009). Social Facilitation of Exploratory Foraging Behavior in Capuchin Monkeys (Cebus Apella). American Journal of Primatology, 71: 419–26.
Ferrari, P.F., Maiolini, C., Addessi, E., Fogassi, L. & Visalberghi, E. (2005). The observation and hearing of eating actions activates motor programs related to eating in macaque monkeys. Behavior Brain Research 161: 95-101.
Fujii, N., Hihara, S. & Iriki A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PLoS ONE 2:e397.
Fujii, N., Hihara, S. & Iriki, S. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience 3:250-60.
Galloway, A.T., Addessi, E., Fragaszy, D. & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchin monkeys (Cebus apella): does it involve behavioral coordination? International Journal of Primatology 26: 175–183.
Guerin, B. & Innes, J. (2009). Social facilitation. Cambridge University Press.
Huguet, P., Galvaing, M.P., Monteil, J.M. & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of Personality and Social Psychology 77: 1011-25.
Leng, Y. & Zhou, X. (2010). Modulation of the brain activity in outcome evaluation by interpersonal relationship : An ERP study. Neuropsychologia, 48, 448-455.
Lonsdorf, E.V., Eberly, L.E. & Pusey, A.E. (2004). Sex differences in learning in chimpanzees. Nature 428:715–716.
Martin, R. (1998). Majority and minority influence using the afterimage paradigm: A series of attempted replications. Journal of Experimental Social Psychology 34: 1-26.
Meunier, M., Monfardini, E. & Boussaoud D. (2007). Learning by observation in rhesus monkeys. Neurobiology of Learning and Memory, 88: 243-8.
Mobbs, D., Yu, R., Meyer, M., Passamonti, L., Seymour, B., Calder, A.J., Schweizer, S., Frith, C.D. & Dalgleish, T. (2010). A Key Role for Similarity in Vicarious Reward. Science, 324 : 2009-2011.
Monfardini, E. (2014). Learning from other’s errors: insights from comparative psychology and human fMRI. Association for Research in Neuroeducation, Caen – France.
Monfardini, E., Gaveau, V., Hadj-Bouziane, F., Boussaoud, D. & Meunier, M. (2012). Social learning as a way to overcome choice-induced preferences? Insights from humans and rhesus macaques. Frontiers of Neurosciences, 6:127. doi:10.3389/fnins.2012.00127.
Monfardini, E., Hadj-Bouziane, F. & Meunier, M. (2014). Model-Observer Similarity, Error Modeling and Social Learning in Rhesus Macaques. PLoS ONE, 9(2) : e89825. doi: 10.1371/journal.pone.0089825.
Perry, S. (2009). Conformism in the food processing techniques of white-faced capuchin monkeys (Cebus capucinus). Animal Cognition, 12, 705-716.
Schunk, D. (1987). Peer models and children’s behavioral change. Review of Educational Research, 57: 149–174.
Schwab, C., Bugnyar, T., Schloegl, C. & Kotrschal, K. (2008). Enhanced social learning between siblings in common ravens, Corvus corax. Animal Behaviour, 75: 501–508.
Visalberghi, E. & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour 60: 69–76.
Visalberghi, E. & Addessi, E. (2001). Acceptance of novel foods in Cebus apella: Do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour 62 : 567– 576.
Visalberghi, E. & Fragaszy, D.M. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour 49: 1089-1095.
| NOM DU PROJET : ISCI3 Phylogénèse de la facilitation sociale de la cognition : du singe à l’homme. |
| Date de début : 2011 | Date de fin : Prévu 2015 |
| R. Psychologie – Neurosciences |
1. Objectifs du projet
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant les neurosciences cognitives, la psychologie sociale, l’éthologie comparée et la neuroendocrinologie. Plus particulièrement, ces travaux portent sur l’influence des interactions sociales sur le fonctionnement cognitif et cérébral individuel. Cela à travers les méthodes scientifiques des neurosciences fondamentales et de la psychologie (Ochsner & Lieberman, 2001).
Le présent axe de recherche s’inscrit dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres. Dans l’approche proposée, le Positionnement Grégaire aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études décrites ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est phylogénétiquement ancien. Étudier cette influence nous permettra donc d’explorer nos hypothèses en termes de cognition sociale et du fonctionnement cérébral qui en est responsable. D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Le but du présent projet de recherche est d’explorer la modulation sociale de la cognition, les mécanismes qui la gouvernent, son évolution au cours de la phylogénèse, ainsi que ses substrats neuronaux. Il vise en particulier à répondre aux questions suivantes :
– Sommes-nous en quelque sorte ‘programmés’ pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (cf. Fig.1, volet Fondamental) ;
– Quelles modulations sociales sont des tendances phylogénétiquement anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes? (cf. Fig.1, volet Fondamental) ;
– Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale? (cf. Fig.1, volet Bases Neurales) ;
– Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions sont abordées via trois approches complémentaires :
– Une approche comportementale comparée confrontant le comportement de l’homme à celui de l’animal mis dans une situation similaire ; (cf. Projets ISCI2 et ISCI3)
– Deux techniques d’imagerie cérébrale humaine : la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) appliquées au macaque et à l’homme (toutes deux permettent une visualisation non invasive de l’activité du cerveau) ; cf Projets ISCI4, ISCI6 et ISCI8)
– L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc. (cf. ISCI7).
D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est simplement la position sociale (« social ranking », en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres (« social comparison »)
Dans ce cadre, le présent projet s’intéresse plus particulièrement au phénomène de facilitation sociale. Ce concept renvoie au fait que la simple présence d’autrui lors de l’accomplissement d’une tâche puisse avoir une influence directe sur nos performances cognitives. Ce phénomène a été largement étudié par la psychologie sociale chez l’homme, et de nombreuses théories ont été avancées pour expliquer ses causes.
Le but de ce projet est de tester l’hypothèse selon laquelle la présence d’autrui lorsqu’on effectue une tâche cognitive peut avoir un effet facilitateur ou inhibiteur aussi bien chez l’homme que chez le primate non-humain, et que cela dépend (i) du niveau de maîtrise de la tâche et (ii) des caractéristiques de l’individu présent.
2. État de l’art
La psychologie sociale a fourni des données expérimentales sur l’influence de la socialité sur le fonctionnement cognitif individuel. Ces études témoignent du fait que la simple présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives, consistant soit à les améliorer, soit à les détériorer (pour une revue récente de la littérature, voir Guerin & Innes, 2009). Le phénomène de modulation sociale concerne une large palette de capacités cognitives : des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements), aux plus complexes comme les capacités attentionnelles (e.g. dans la tâche de Stroop, Huguet et al., 1999), la prise de décision (Asch, 1956 ; Martin, 1998), ou l’apprentissage social de règles (Bounoua et al., 2009). En résumé, des situations sociales relativement simples peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales ou cognitives.
Les effets d’influence sociale ne sont pas une prérogative humaine. Des phénomènes de facilitation sociale ont aussi été démontrés chez les primates non-humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire. Par exemple, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001 ; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Au niveau cérébral, une seule série d’études a jusqu’à présent donné quelques éléments pour répondre à cette question. Les auteurs (Fujii et al., 2007 ; 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone.
Comme nous l’avons précédemment dit, l’apprentissage social est également une composante fondamentale de la cognition humaine. Apprendre par observation d’autrui est vital pour l’homme (Castro & Toro, 2004). Sans cet apprentissage, chaque enfant devrait réapprendre chaque comportement laborieusement par lui-même et aucune transmission de savoir-faire ne serait possible. Selon Bandura (1965), ce type d’apprentissage est caractérisé par quatre étapes: l’attention, le maintien en mémoire, la reproduction du geste, et la motivation. La première étape, l’attention, est un processus crucial et peut être fortement influencé par les caractéristiques du modèle. Il a été démontré que des caractéristiques telles que l’attractivité, la similitude et la compétence perçue améliorent l’efficacité de l’apprentissage par observation (Brewer & Wann, 1998). En particulier, la similitude entre acteur et observateur serait la clé d’une transmission sociale des connaissances efficace. Adultes comme enfants savent en effet bénéficier de l’observation d’individus dont les performances sont légèrement supérieures quand ils s’agit d’apprendre des associations stimulus-récompense, mais au contraire ne bénéficient pas de l’observation d’individus perçus comme trop experts (Bounoua et al., 2009). Des récentes données de neurosciences corroborent ces données, en montrant une modulation des mécanismes cérébraux responsables de l’apprentissage social selon le dégrée de similarité et/ou affinité perçu entre modèle et apprenti (Mobbs et al., 2009 ; Leng & Zhou, 2010).
3. Aléas, incertitudes scientifiques, verrous technologiques
L’étude neuroscientifique de la modulation sociale de la cognition et, de façon plus générale, de la sphère sociale de notre existence, présente un grand intérêt mais aussi des défis techniques considérables.
Le défi essentiel de ce domaine est le contrôle d’un grand nombre de variables qui interviennent dans nos rapports sociaux et qui influencent notre « positionnement grégaire ». La complexité de ce domaine de recherche implique la multiplication des variables expérimentales à inclure dans nos protocoles expérimentaux et, par conséquent, des coûts considérables en termes de temps et de prix d’examens (notamment pour ce qui concerne les études neuroscientifiques d’imagerie). Cependant, les collaborations scientifiques sur lesquelles ces études reposent nous permettent de faire face aux problèmes liés à l’originalité de ce domaine d’étude. Ainsi, les résultats de ces études ont déjà fait l’objet de plusieurs publications dans des journaux scientifiques internationaux (Plos One, NeuroImage, Frontiers In Neurosciences, etc) et positionnent le Pôle de Recherche de l’IME dans le panorama scientifique international aussi bien dans le domaine de la Psychologie Cognitive que des Neurosciences.
L’étude ISCI3 implique l’utilisation d’un modèle animal, le primate non-humain. Ceci rentre dans le cadre d’une approche comparative finalisée à isoler les composantes fondamentales des comportements humains, qui s’expriment indépendamment de la culture et du langage. En particulier, le choix du modèle primate est motivé par l’intérêt que nous portons aux hiérarchies sociales et au positionnement grégaire. Or, les singes macaques passent la majeure partie de leur vie éveillée en présence de leurs congénères, et les rapports qu’ils instaurent au sein d’un groupe sont gérés par de fortes hiérarchies sociales. L’utilisation d’un modèle animal comporte des coûts et des aléas tout à fait différents de ceux auxquels on fait face pour les études chez l’homme. Tout d’abord, l’hébergement des animaux requiert du personnel et des infrastructures très coûteuses. Ensuite, la réalisation de protocoles expérimentaux impliquant des tâches cognitives requiert l’entrainement des animaux, qui se fait de façon progressive souvent sur des longues périodes (1 à 6 mois). Cet entrainement, qui comporte des travers les méthodes de conditionnement opérant bien connues en éthologie et psychologie comparative, demande des compétences particulières de la part du personnel impliqué dans l’étude. Le projet ISCI3 a été mené avec succès en collaboration avec le Centre de Recherche en Neurosciences de Lyon, qui possède une animalerie primate complètement rénovée et pouvant héberger un nombre important d’animaux. Le personnel INSERM qui y travaille a assuré le soin des animaux et leur hébergement conformément aux directives européennes pendant tout le déroulement du projet.
4. Travaux R&D réalisés, démarche expérimentale
Pour ce projet, nous avons développé un système expérimental impliquant des ordinateurs à écran tactile et des distributeurs de récompense (Fig.2), et égalisé le niveau de difficulté des tâches pour les deux espèces testées, de telle sorte à pouvoir comparer les données issues des deux espèces. Le protocole expérimental conçu est de type factoriel 2x3x2, avec comme premier facteur le niveau de maîtrise de la tâche (tâche maîtrisée ; tâche non maîtrisée) comme deuxième facteur la situation sociale (isolation sociale ; présence d’un congénère familier ; présence d’un congénère inconnu), et comme troisième facteur ce que l’individu présent fait pendant l’exécution de la tâche (« audience », i.e. simple présence d’un individu passif ; « coaction », i.e. l’autre individu présent effectue lui aussi la tâche au même temps).
Nous avons actuellement finalisé l’expérience chez le singe ainsi que chez l’homme. L’expérience chez l’homme a été le sujet de stage pour un étudiant en Médecine et de trois étudiantes en Psychologie Expérimentale.

Figure 2. Système expérimental utilisé pour les expériences chez les primates humains (à droite) et non-humains (à gauche) de l’étude comparative ISCI3.
Les résultats chez les singes (n =7) suggèrent que ni le degré de maîtrise de la tâche ni l’activité du compagnon ne modulent l’effet de la présence d’autrui, qui est dans tous les cas extrêmement facilitatrice chez les animaux.
5. Indicateurs de R&D
Les résultats de cette étude ont été présenté lors de deux conférences (Monfardini et al., 2014 ; Monfardini, 2013) et un article scientifique est en cours de préparation (Monfardini & Meunier, en cours).
6. Acquisition de connaissances
Les résultats de cette étude tendent à montrer qu’en présence d’autrui, les performances sont améliorées aussi bien dans le cas d’une tâche maitrisée que dans une tâche non maîtrisée, et aussi bien quand l’autre individu présent travaille que quand il est passif. En revanche, la qualité de la relation entre les deux acteurs sociaux a une influence sur les performances cognitives : travailler en présence d’un individu non familier (dont le rang social est donc inconnu) inhiberait fortement les performances au point d’annuler toute sorte de phénomène de facilitation sociale chez le singe.
D’un point de vue applicatif, les résultats de cette recherche ouvrent la voie à des réflexions théoriques et à des travaux empirique d’intérêt sociétal majeur. Il existe aujourd’hui une forte demande d’optimisation des performances de la part des individus sains (Lynch et al., 2011 ; Husain et Mehta, 2011; Frenguelli, 2013). Dans une enquête informelle effectuée par l’éminent journal Nature en 2009 auprès de son lectorat, environ un quart des lecteurs de plus de 50 ans, chercheurs et professeurs d’université, reconnaissaient avoir eu recours à des psychostimulants comme la Ritaline ou le Modafinil. En France, les ventes de Ritaline ont augmenté de 70% entre 2008 et 2013 en France selon Le Parisien du 29/05/2013 et la consommation par des sujets sains contribue sans aucun doute à ce phénomène. Cette évolution inquiétante souligne la nécessité de trouver des alternatives non pharmacologiques pour répondre à cet enjeu sociétal majeur. Le fait que la simple présence d’autrui module d’une façon majeure nos capacités et habilités cognitives ouvre des pistes dans cette direction : est-ce la dimension sociales capable d’améliorer la concentration, autant qu’une drogue qui augmente la noradrénaline dans le cerveau ?
Les résultats de cette étude, ensemble avec d’autres qui en découleront feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach, et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
7. Ressources humaines
| Nom | Fonction dans le projet | Nbre heures/jours consacrées à la Phase 1 | Nbre heures/jours consacrées à la Phase 2 | Nbre heures/jours consacrées à la Phase 3 | Nb total d’heures/jours affectées au projet |
| Elisabetta Monfardini | Chef de projet |
Recherche bibliographique, recherche de collaboration, demande d’autorisations, construction du protocole (2011) |
Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | Traitement des résultats et écriture de l’article (2014-2015) | 86 heures |
| Fradin Jacques | Directeur de recherche | 29 heures |
8. Références bibliographiques
Asch, S.E. (1956). Studies of independence and conformity: I. A minority of one against a unanimous majority. Psychological Monographs 70(9, Whole No. 416).
Bandura, A. (1965). Influence of models’ reinforcement contingencies on the acquisition of imitative responses. Journal of Personnality and Social Psychology, 1, 589-595.
Bounoua, L., Cury, F., Monfardini, E., Meunier, M., Boussaoud, D., Brovelli, A. & Huguet, P. (2009). Learning by observation: The role of social comparison and related ability inferences. 16th Annual ESCOP conference, Jagiellonian University, Krakow.
Brewer, K.R., Wann, D. L. (1998). Observational learning effectiveness as a function of model characteristics: Investigating the importance of social power. Social Behavior and Personality, 26(1), 1-10.
Castro, L. & Toro, M. (2004). The evolution of culture: From primate social learning to human culture. Proceedings of the National Academy of Sciences, 101: 10235-10240.
Dindo, M., Whiten, A. & de Waal, F. B. M. (2009). Social Facilitation of Exploratory Foraging Behavior in Capuchin Monkeys (Cebus Apella). American Journal of Primatology, 71: 419–26.
Ferrari, P.F., Maiolini, C., Addessi, E., Fogassi, L. & Visalberghi, E. (2005). The observation and hearing of eating actions activates motor programs related to eating in macaque monkeys. Behavior Brain Research 161: 95-101.
Frenguelli, B.G. (2013). Cognitive enhancers: Molecules, mechanisms and minds. Neuropharmacology, 64: 1.
Fujii, N., Hihara, S. & Iriki A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PLoS ONE 2:e397.
Fujii, N., Hihara, S. & Iriki, S. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience 3:250-60.
Galloway, A.T., Addessi, E., Fragaszy, D. & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchin monkeys (Cebus apella): does it involve behavioral coordination? International Journal of Primatology 26: 175–183.
Guerin, B. & Innes, J. (2009). Social facilitation. Cambridge University Press.
Huguet, P., Galvaing, M.P., Monteil, J.M. & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of Personality and Social Psychology 77: 1011-25.
Husain, M. & Mehta, M.A. (2011). Cognitive enhancement by drugs in health and disease. Trends in Cognitive Sciences, 15:28-36.
Leng, Y. & Zhou, X. (2010). Modulation of the brain activity in outcome evaluation by interpersonal relationship : An ERP study. Neuropsychologia, 48, 448-455.
Lynch G., Palmer LC., Gall C M. (2011). The likelihood of cognitive enhancement. Pharmacology Biochemistry and Behavior, 99(2): 116-129.
Martin, R. (1998). Majority and minority influence using the afterimage paradigm: A series of attempted replications. Journal of Experimental Social Psychology 34: 1-26.
Meunier, M., Monfardini, E. & Boussaoud D. (2007). Learning by observation in rhesus monkeys. Neurobiology of Learning and Memory, 88: 243-8.
Mobbs, D., Yu, R., Meyer, M., Passamonti, L., Seymour, B., Calder, A.J., Schweizer, S., Frith, C.D. & Dalgleish, T. (2010). A Key Role for Similarity in Vicarious Reward. Science, 324 : 2009-2011.
Monfardini, E. (2013). Social facilitation in monkeys. Talk. SONA International Conference, Rabat – Maroc.
Monfardini, E., Gaveau, V., Hadj-Bouziane, F., Boussaoud, D. & Meunier, M. (2012). Social learning as a way to overcome choice-induced preferences? Insights from humans and rhesus macaques. Frontiers of Neurosciences, 6, 127. doi:10.3389/fnins.2012.00127.
Monfardini, E. & Meunier, M. (en préparation). The social influence on cognition: A comparative study.
Monfardini, E., Reynaud, A., Guedj, C., Hynaux, C., Gardechaux, G., Farnè, A., Hadj-Bouziane, F. & Meunier, M. (2014). Social facilitation of cognition in macaque monkeys. CogEvo Workshop on Cognition and Evolution, Rovereto – Italy.
Visalberghi, E. & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour 60: 69–76.
Visalberghi, E. & Addessi, E. (2001). Acceptance of novel foods in Cebus apella: Do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour 62 : 567– 576.
Visalberghi, E. & Fragaszy, D.M. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour 49: 1089-1095.
| NOM DU PROJET : ISCI4-Bases neuronales de la facilitation sociale : Mesure du métabolisme cérébral par tomographie par émission de positrons ([18F]-FDG TEP) chez le singe macaque rhésus |
| Date de début : 2011 | Date de fin : Prévu 2015 |
| R. Psychologie – Neurosciences |
1. Objectifs du projet
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant les neurosciences cognitives, la psychologie sociale, l’éthologie comparée et la neuroendocrinologie. Plus particulièrement, ces travaux portent sur l’influence des interactions sociales sur le fonctionnement cognitif et cérébral individuel. Cela à travers les méthodes scientifiques des neurosciences fondamentales et de la psychologie (Ochsner & Lieberman, 2001).
Le présent axe de recherche s’inscrit dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres. Dans l’approche proposée, le Positionnement Grégaire aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études décrites ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est phylogénétiquement ancien. Étudier cette influence nous permettra donc d’explorer nos hypothèses en termes de cognition sociale et du fonctionnement cérébral qui en est responsable. D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Le but du présent projet de recherche est d’explorer la modulation sociale de la cognition, les mécanismes qui la gouvernent, son évolution au cours de la phylogénèse, ainsi que ses substrats neuronaux. Il vise en particulier à répondre aux questions suivantes :
– Sommes-nous en quelque sorte ‘programmés’ pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (Fig.1, volet Fondamental) ;
– Quelles modulations sociales sont des tendances phylogénétiquement anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes? (cf. Fig.1, volet Fondamental) ;
– Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale? (cf. Fig.1, volet Bases Neurales) ;
– Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions sont abordées via trois approches complémentaires :
– Une approche comportementale comparée confrontant le comportement de l’homme à celui de l’animal mis dans une situation similaire ; (cf. Projets ISCI2 et ISCI3)
– Deux techniques d’imagerie cérébrale humaine : la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) appliquées au macaque et à l’homme (toutes deux permettent une visualisation non invasive de l’activité du cerveau) ; cf. Projets ISCI4, ISCI6 et ISCI8)
– L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc. (cf. ISCI7).
D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est simplement la position sociale (« social ranking », en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres (« social comparison »)
Dans ce cadre, le présent projet s’intéresse plus particulièrement aux bases neuronales sous-jacentes au phénomène de facilitation sociale. Ce concept renvoie au fait que la simple présence d’autrui lors de l’accomplissement d’une tâche puisse avoir une influence directe sur nos performances cognitives. Ce phénomène a été largement étudié par la psychologie sociale chez l’homme, et de nombreuses théories ont été avancées pour expliquer ses causes.
2. État de l’art
La psychologie sociale a fourni des données expérimentales sur l’influence de la socialité sur le fonctionnement cognitif individuel. Ces études témoignent du fait que la simple présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives, consistant soit à les améliorer, soit à les détériorer (pour une revue récente de la littérature, voir Guerin & Innes, 2009). Le phénomène de modulation sociale concerne une large palette de capacités cognitives : des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements), aux plus complexes comme les capacités attentionnelles (e.g. dans la tâche de Stroop, Huguet et al., 1999), la prise de décision (Asch, 1956 ; Martin, 1998 ), ou l’apprentissage social de règles (Bounoua et al., 2009). En résumé, des situations sociales relativement simples peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales ou cognitives.
Les effets d’influence sociale ne sont pas une prérogative humaine. Des phénomènes de facilitation sociale ont aussi été démontrés chez les primates non-humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire. Par exemple, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Au niveau cérébral, une seule série d’études a jusqu’à présent donné quelques éléments pour répondre à cette question. Les auteurs (Fujii et al., 2007 ; 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone.
Comme nous l’avons précédemment dit, l’apprentissage social est également une composante fondamentale de la cognition humaine. Apprendre par observation d’autrui est vital pour l’homme (Castro & Toro, 2004). Sans cet apprentissage, chaque enfant devrait réapprendre chaque comportement laborieusement par lui-même et aucune transmission de savoir-faire ne serait possible. Selon Bandura (1965), ce type d’apprentissage est caractérisé par quatre étapes: l’attention, le maintien en mémoire, la reproduction du geste, et la motivation. La première étape, l’attention, est un processus crucial et peut être fortement influencé par les caractéristiques du modèle. Il a été démontré que des caractéristiques telles que l’attractivité, la similitude et la compétence perçue améliorent l’efficacité de l’apprentissage par observation (Brewer & Wann, 1998). En particulier, la similitude entre acteur et observateur serait la clé d’une transmission sociale des connaissances efficace. Adultes comme enfants savent en effet bénéficier de l’observation d’individus dont les performances sont légèrement supérieures quand ils s’agit d’apprendre des associations stimulus-récompense, mais au contraire ne bénéficient pas de l’observation d’individus perçus comme trop experts (Bounoua et al., 2009). Des récentes données de neurosciences corroborent ces données, en montrant une modulation des mécanismes cérébraux responsables de l’apprentissage social selon le dégrée de similarité et/ou affinité perçu entre modèle et apprenti (Mobbs et al., 2009 ; Leng & Zhou, 2010).
Dans ce cadre, le présent projet a eu pour objectif de mettre en place un protocole d’imagerie qui soit compatible avec une situation d’interaction sociale chez le singe, et impliquant notamment un contexte d’apprentissage par facilitation sociale.
3. Aléas, incertitudes scientifiques, verrous technologiques
L’étude neuroscientifique de la modulation sociale de la cognition et, plus en général, de la sphère sociale de notre existence, présente un grand intérêt mais aussi des défis techniques considérables. Premier élément, le contrôle d’un grand nombre de variables qui interviennent dans nos rapports sociaux et qui influencent notre « positionnement grégaire ». La complexité de ce domaine de recherche implique la multiplication des variables expérimentales à inclure dans nos protocoles expérimentaux et, par conséquent, des coûts considérables en termes de temps et de prix d’examens (notamment pour ce qui concerne les études neuroscientifiques d’imagerie). Cependant, les collaborations scientifiques sur lesquelles ces études reposent, ainsi que les publications scientifiques nationales et internationales qui en découlent, nous permettent de faire face aux problèmes liés à l’originalité de ce domaine d’étude. En effet, les résultats de ces études ont déjà fait l’objet de plusieurs publications dans des journaux scientifiques internationaux (Plos One, NeuroImage, Frontiers In Neurosciences, etc.) et positionnent le Pôle de Recherche de l’IME dans le panorama scientifique international aussi bien dans le domaine de la Psychologie Cognitive que des Neurosciences.
Pour cette étude, nous avons été confrontés au fait que l’IRMf, combinée aux méthodes invasives possibles chez le singe est parfaitement adaptée à l’étude de la cognition chez le sujet isolé, mais son apport est forcément limité quand il s’agit d’aborder les interactions sociales. En effet, la plupart des études en imagerie cérébrale sur la cognition sociale ont jusqu’à présent mis en évidence les activations cérébrales relatives à la présentation de stimuli de validité écologique incertaine ou douteuse. Pour dépasser cette limite, nous avons proposé d’appliquer au singe un type particulier de protocole utilisant la technique d’imagerie de la TEP. Nous utilisons le [18F]-fluorodeoxyglucose ([18F]-FDG), qui est un marqueur du métabolisme cérébral du glucose, et sa visualisation par la TEP pour définir la réponse neuronale liée à la modulation sociale pendant une tâche. L’avantage de cette approche en TEP est la possibilité d’étudier la réponse cérébrale à l’engagement dans une interaction sociale ’écologique’ (eg. Rilling et al, 2004). L’animal peut en effet être placé dans une situation sociale naturelle (phase dite ‘d’activation’) et être scanné seulement ensuite, à l’état vigile ou sous anesthésie.
Au cours des l’année 2011/2012 nous avons mis au point cette technique, inexplorée en France malgré son intérêt dans l’étude des bases neuronales de la cognition sociale, et finalisé la procédure de traitement des données brutes.
4. Travaux R&D réalisés, démarche expérimentale
Pour cette étude, nous avons donc mis au point une technique d’imagerie (IRMf) permettant l’étude des bases neuronales de la cognition sociale, et finalisé la procédure de traitement des données brutes. Les images statiques TEP [18F]FDG de radioactivité brute sont en effet normalisées par la dose injectée et par le poids du singe, pour former une image paramétrique dite Standardized Uptake Value (SUV). Cette image reflète la consommation métabolique régionale de glucose (CRMgl). Les images individuelles d’un sujet sont recalées sur son IRM anatomique T1, puis normalisées dans un espace stéréotaxique de cerveau de singe standard. A l’issue de cette normalisation spatiale, nous pouvons effectuer une analyse statistique voxel à voxel du plan factoriel mis en œuvre dans cette expérience. La taille de notre échantillon et la connaissance a priori de l’erreur typique de mesure prédisent que cette analyse statistique sera réalisée avec une puissance de 0.9 pour la détection de modification de CMRgl de 20%.
Nous avons ainsi pu effectuer une première expérience chez le singe macaque rhésus. Cette étude a pour but d’explorer les aires cérébrales responsables de l’effet facilitateur de la présence sociale sur les performances cognitives individuelles, sujet d’étude jusqu’à maintenant inexploré au sein de la communauté neuroscientifique. La tâche utilisée est une simple tâche d’association stimulus-récompense (Meunier et al, 2007). L’animal est placé face à un écran tactile au centre duquel apparaissent des images, et il doit toucher l’image positive pour obtenir une récompense. Chaque session consiste à injecter le [18F]-FDG à l’un des membres de la dyade qui est ensuite exposé à l’une des trois conditions expérimentales suivantes (Fig.2; temps d’exposition: 30 minutes) :
1) tâche cognitive en présence sociale
2) tâche cognitive en isolation sociale
3) condition contrôle (singe passif en isolation social)



Figure 2. Conditions expérimentales du projet ISCI4. (1) condition Présence Sociale ; (2) condition Isolation ; (3) condition de contrôle.
Après la période de d’absorption du [18F]-FDG (30 minutes), le singe est anesthésié (Zoletil 100) et transporté au Cermep pour l’examen TEP. L’analyse statistique des données prétraitées permet de corréler les performances des individus dans la tâche au sein des trois conditions expérimentales. Parce que le ‘statut social’ est connu pour affecter les performances cognitives chez les singes (Drea & Wallen, 1999), cette variable est contrôlée aux cours de l’expérience. Une batterie de tests comportementaux est en effet utilisée pour identifier le dominant et le subordonné au sein de chaque dyade.
Les résultats de la totalité des sessions expérimentales (n = 27) démontrent que le réseau attentionnel fronto-pariétal (i.e., prémoteur dorsal, pariétal supérieur) joue un rôle dans la facilitation sociale (Fig. 3).
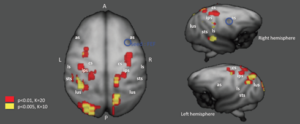
Figure 3. Résultats de l’étude 4. Le réseau cérébral attentionnel fronto-parietal est plus activé dans la condition de présence sociale. Ceci ce traduit en une amélioration de sa performance, en plaidant pour la thèse de la genèse attentionnelle du phénomène de facilitation sociale (Huguet et al, 1999).
5. Indicateurs de R&D
Les résultats de cette étude ont été présentés dans plusieurs congrès internationaux (Monfardini, 2014a ; Monfardini, 2014b ; Monfardini , 2013 ; Monfardini et al., 2013). Cette étude a été récompensée par l’Université de Chicago comme meilleure étude dans le domaine de la cognition animale pour le congrès de la Society for Social Neuroscience. L’article décrivant les résultats de cette étude est en cours de finalisation et sa publication dans un journal international est le prochain objectif pour 2014.
6. Acquisition de connaissances
Selon les résultats de cette étude, la simple présence d’autrui lors de la réalisation d’une tâche serait capable de moduler l’activité cérébrale au sein du réseau fronto-pariétal de l’attention. Les autres changeraient notre capacité à prêter attention à une tâche, et ceci en agissant sur nos mécanismes cérébraux « de base ». Ce résultat est de grand intérêt pour au moins deux raisons : 1) d’un point de vue fondamental, suggère que le cerveau possède des propriétés « sociales » intrinsèques, qu’il est fondamental de connaître afin de comprendre son fonctionnement; 2) d’un point de vue plus appliqué, incite à tenir en compte de l’aspect social quand on veut optimiser nos performances attentionnelles. L’attention est impliquée dans toutes nos activités intellectuelles. Elle nous permet soit d’être globalement vigilants, soit de focaliser notre concentration sur un point précis, soit au contraire de répartir nos ressources entre plusieurs activités. Sans attention, les fonctions cognitives supérieures ne peuvent plus fonctionner correctement. Isoler les conditions qui favorisent nos capacités attentionnelles est donc un enjeu majeur, notamment en contexte professionnel ou pédagogique. Il existe aujourd’hui une forte demande d’optimisation des performances de la part des entreprises, comme de la part des individus. Les salariés gaspilleraient plus de deux heures de travail par jour du fait d’interruptions diverses, entrainant 588 milliards de dollars de pertes par an pour les seuls USA
(http://iorgforum.org/wpcontent/uploads/2011/06/CostOfNotPayingAttention.BasexReport.pdf). Apprendre à leurs employés à maîtriser leur attention est donc un véritable enjeu économique pour les entreprises. Du côté des individus, l’extrême compétitivité du marché du travail conduit de nombreux professionnels à chercher de l’aide pour se maintenir dans un emploi ou accéder à une promotion. Certains ont recours aux psychostimulants, un phénomène qui explique en partie l’augmentation des ventes des ‘dopants cognitifs’ comme la Ritaline (70% entre 2008 et 2013 en France selon Le Parisien, 29 mai 2013), une évolution inquiétante pour un médicament dont l’action sur le cerveau reste mal connue. Les résultats de la présente étude permettent de suggérer des pistes d’exploration neuroscientifique capables de proposer des solutions non médicamenteuses basées sur le fonctionnement cérébral en situation sociale pour optimiser notre attention. Des études ultérieures sont prévues pour 2014 dans ce domaine.
7. Ressources humaines
| Nom | Fonction dans le projet | Nbre heures/jours consacrées à la Phase 1 | Nbre heures/jours consacrées à la Phase 2 | Nbre heures/jours consacrées à la Phase 3 | Nb total d’heures/jours affectées au projet |
| Elisabetta Monfardini | Chef de projet |
Recherche bibliographique, recherche de collaboration, demande d’autorisations, construction du protocole (2011) |
Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | Traitement des résultats et écriture de l’article (2014-2015) | 346 heures |
| Jacques Fradin | Directeur de recherche |
Recherche bibliographique, recherche de collaboration, demande d’autorisations, construction du protocole (2011) |
Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | Traitement des résultats et écriture de l’article (2014-2015) | 30 heures |
8. Références bibliographiques
Asch, S.E. (1956). Studies of independence and conformity: I. A minority of one against a unanimous majority. Psychological Monographs 70(9, Whole No. 416).
Bounoua, L., Cury, F., Monfardini, E., Meunier, M., Boussaoud, D., Brovelli, A. & Huguet, P. (2009). Learning by observation: The role of social comparison and related ability inferences. 16th Annual ESCOP conference, Jagiellonian University, Krakow.
Dindo, M., Whiten, A. & de Waal, F. B. M. (2009). Social Facilitation of Exploratory Foraging Behavior in Capuchin Monkeys (Cebus Apella). American Journal of Primatology, 71, 419–26.
Drea, C.M. & Wallen, K. (1999). Low status monkeys “play dumb” when learning in mixed social groups. Proceedings of the National Academy of Sciences, 96: 12965-12969.
Ferrari, P.F., Maiolini, C., Addessi, E., Fogassi, L. & Visalberghi, E. (2005). The observation and hearing of eating actions activates motor programs related to eating in macaque monkeys. Behavior Brain Research 161, 95-101.
Fujii, N., Hihara, S. & Iriki A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PLoS ONE 2:e397.
Fujii, N., Hihara, S. & Iriki, S. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience 3, 250-60.
Galloway, A.T., Addessi, E., Fragaszy, D. & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchin monkeys (Cebus apella): does it involve behavioral coordination? International Journal of Primatology 26, 175–183.
Guerin, B. & Innes, J. (2009). Social facilitation. Cambridge University Press.
Huguet, P., Galvaing, M.P., Monteil, J.M. & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of Personality and Social Psychology 77, 1011-25.
Martin, R. (1998). Majority and minority influence using the afterimage paradigm: A series of attempted replications. Journal of Experimental Social Psychology 34: 1-26.
Meunier, M., Monfardini, E. & Boussaoud D. (2007). Learning by observation in rhesus monkeys. Neurobiology of Learning and Memory, 88, 243-8.
Monfardini, E. (2013). Social facilitation in monkeys. Talk. SONA International Conference, Rabat – Maroc.
Monfardini, E. (2014). Learning from other’s errors: insights from comparative psychology and human fMRI. Association for Research in Neuroeducation, Caen – France.
Monfardini, E. (2014). Non-invasive neuroimaging in the macaque monkey. EFOR meeting, Paris- France.
Monfardini, E., Redouté, J., Hadj-Bouziane, F., Hynaux, C., Fradin, J., Huguet, P., Costes, N. & Meunier, M. (under revision). Others’ sheer presence boosts brain activity in the attention (but not the motivation) network. Cerebral Cortex.
Monfardini, E., Redouté, J., Hynaux, C., Le Bars, D., Lavenne, F., Fonlup, P., Costes, N. & Meunier, M. (2012). Neural basis of social facilitation in monkeys using FDG-PET. Poster. Society for Neuroscience Meeting, New Orleans – Louisiana.
Monfardini, E., Redouté, J., Hynaux, C., Le Bars, D., Lavenne, F., Fonlup, P., Costes, N. & Meunier, M. (2012). Social facilitation in monkeys. Poster. Society for Social Neuroscience Meeting, New Orleans – Louisiana (Best Poster Presentation awarded).
Monfardini, E., Redouté, J., Hynaux, C., Le Bars, D., Lavenne, F., Fonlup, P., Costes, N. & Meunier, M. (2013). Neural basis of social facilitation in monkeys. Poster. Third International Symposium on « The Biology of Decision Making » (SBDM 2013), Paris – France.
Rilling, J.K., Winslow, J.T. & Kilts, C.D. (2004). The neural correlates of mate competition in dominant male rhesus macaques. Biological Psychiatry 56: 364-75.
Visalberghi, E. & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour 60: 69–76.
Visalberghi, E. & Addessi, E. (2001). Acceptance of novel foods in Cebus apella: Do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour 62 : 567– 576.
Visalberghi, E. & Fragaszy, D.M. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour 49: 1089-1095.
| NOM DU PROJET : ISCI6 Mécanismes cérébraux de l’apprentissage social chez l’homme : une étude en IRMf. |
| Date de début : 2013 | Date de fin : 2013 |
| R. Psychologie – Neurosciences |
1. Objectifs du projet
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant les neurosciences cognitives, la psychologie sociale, l’éthologie comparée et la neuroendocrinologie. Plus particulièrement, ces travaux portent sur l’influence des interactions sociales sur le fonctionnement cognitif et cérébral individuel. Cela à travers les méthodes scientifiques des neurosciences fondamentales et de la psychologie (Ochsner & Lieberman, 2001).
Le présent axe de recherche s’inscrit dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres. Dans l’approche proposée, le Positionnement Grégaire aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études décrites ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est phylogénétiquement ancien. Étudier cette influence nous permettra donc d’explorer nos hypothèses en termes de cognition sociale et du fonctionnement cérébral qui en est responsable. D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Le but du présent projet de recherche est d’explorer la modulation sociale de la cognition, les mécanismes qui la gouvernent, son évolution au cours de la phylogénèse, ainsi que ses substrats neuronaux. Il vise en particulier à répondre aux questions suivantes :
– Sommes-nous en quelque sorte ‘programmés’ pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (cf. Fig.1, volet Fondamental) ;
– Quelles modulations sociales sont des tendances phylogénétiquement anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes? (cf. Fig.1, volet Fondamental) ;
– Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale? (cf. Fig.1, volet Bases Neurales) ;
– Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions sont abordées via trois approches complémentaires :
– Une approche comportementale comparée confrontant le comportement de l’homme à celui de l’animal mis dans une situation similaire ; (cf. Projets ISCI2 et ISCI3)
– Deux techniques d’imagerie cérébrale humaine : la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) appliquées au macaque et à l’homme (toutes deux permettent une visualisation non invasive de l’activité du cerveau) ; cf. Projets ISCI4, ISCI6 et ISCI8)
– L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc. (cf. ISCI7).
D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est simplement la position sociale (« social ranking », en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres (« social comparison »)
Dans ce cadre, le présent projet s’intéresse plus particulièrement aux bases neuronales sous-jacentes à l’apprentissage social chez l’homme.
2. État de l’art
La psychologie sociale a fourni des données expérimentales sur l’influence de la socialité sur le fonctionnement cognitif individuel. Ces études témoignent du fait que la simple présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives, consistant soit à les améliorer, soit à les détériorer (pour une revue récente de la littérature, voir Guerin & Innes, 2009). Le phénomène de modulation sociale concerne une large palette de capacités cognitives : des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements), aux plus complexes comme les capacités attentionnelles (e.g. dans la tâche de Stroop, Huguet et al., 1999), la prise de décision (Asch, 1956 ; Martin, 1998), ou l’apprentissage social de règles (Bounoua et al., 2009). En résumé, des situations sociales relativement simples peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales ou cognitives.
Les effets d’influence sociale ne sont pas une prérogative humaine. Des phénomènes de facilitation sociale ont aussi été démontrés chez les primates non-humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire. Par exemple, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001 ; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Au niveau cérébral, une seule série d’études a jusqu’à présent donné quelques éléments pour répondre à cette question. Les auteurs (Fujii et al., 2007 ; 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone.
Comme nous l’avons précédemment dit, l’apprentissage social est également une composante fondamentale de la cognition humaine. Apprendre par observation d’autrui est vital pour l’homme (Castro & Toro, 2004). Sans cet apprentissage, chaque enfant devrait réapprendre chaque comportement laborieusement par lui-même et aucune transmission de savoir-faire ne serait possible. Selon Bandura (1965), ce type d’apprentissage est caractérisé par quatre étapes: l’attention, le maintien en mémoire, la reproduction du geste, et la motivation. La première étape, l’attention, est un processus crucial et peut être fortement influencé par les caractéristiques du modèle. Il a été démontré que des caractéristiques telles que l’attractivité, la similitude et la compétence perçue améliorent l’efficacité de l’apprentissage par observation (Brewer & Wann, 1998). En particulier, la similitude entre acteur et observateur serait la clé d’une transmission sociale des connaissances efficace. Adultes comme enfants savent en effet bénéficier de l’observation d’individus dont les performances sont légèrement supérieures quand ils s’agit d’apprendre des associations stimulus-récompense, mais au contraire ne bénéficient pas de l’observation d’individus perçus comme trop experts (Bounoua et al., 2009). Des récentes données de neurosciences corroborent ces données, en montrant une modulation des mécanismes cérébraux responsables de l’apprentissage social selon le dégrée de similarité et/ou affinité perçu entre modèle et apprenti (Mobbs et al., 2009 ; Leng & Zhou, 2010).
Dans ce cadre, le but de la présente étude était de comparer les bases cérébrales de l’apprentissage social avec celles de l’apprentissage par essai et erreur.
3. Aléas, incertitudes scientifiques, verrous technologiques
L’étude neuroscientifique de la modulation sociale de la cognition et, plus en général, de la sphère sociale de notre existence, présente un grand intérêt mais aussi des défis techniques considérables. Premier élément, le contrôle d’un grand nombre de variables qui interviennent dans nos rapports sociaux et qui influencent notre « positionnement grégaire ». La complexité de ce domaine de recherche implique la multiplication des variables expérimentales à inclure dans nos protocoles expérimentaux et, par conséquent, des coûts considérables en termes de temps et de prix d’examens (notamment pour ce qui concerne les études neuroscientifiques d’imagerie). Cependant, les collaborations scientifiques sur lesquelles ces études reposent, ainsi que les publications scientifiques nationales et internationales qui en découlent, nous permettent de faire face aux problèmes liés à l’originalité de ce domaine d’étude. En effet, les résultats de ces études ont déjà fait l’objet de plusieurs publications dans des journaux scientifiques internationaux (Plos One, NeuroImage, Frontiers In Neurosciences, etc.) et positionnent le Pôle de Recherche de l’IME dans le panorama scientifique international aussi bien dans le domaine de la Psychologie Cognitive que des Neurosciences.
L’étude ISCI6 a été menée dans le cadre d’une collaboration internationale, avec les Drs Bruno Wicker et Andrea Brovelli de l’Institut de Neurosciences de la Timone – CNRS (Marseille), le Dr Driss Boussaoud de l’Institut de Neurosciences des Systèmes – INSERM (Marseille), ainsi que les Drs Christian Keysers et Valeria Gazzola du Netherlands Institute for Neuroscience, Royal Netherlands Academy of Arts and Sciences (Amsterdam, The Netherlands). Ceci a permis de supporter les coûts importants en terme de réalisation de l’expérience et de publication de l’article scientifique. L’expérience d’imagerie, en effet, a été réalisée au préalable à l’Université de Groningen (Hollande), mais les données ont été analysées courant 2013 par Elisabetta Monfardini.
4. Travaux R&D réalisés, démarche expérimentale
Le but de la présente étude était de comparer les bases cérébrales de l’apprentissage social avec celles de l’apprentissage par essai et erreur. Nous avons choisi la résonance magnétique fonctionnelle (IRMf), une technique d’imagerie qui permet d’obtenir une cartographie très précise des activations cérébrales liées à la réalisation d’une tâche. Les résultats que nous avons obtenus, qui ont été publiés dans la revue PlosOne en 2013, suggèrent que le traitement de l’information pendant l’apprentissage social et l’apprentissage individuel engage des réseaux cérébrales communs (Figure 2). Ces réseaux incluent le système fronto-parietal, la boucle associative fronto-striatale, ainsi que le système de neurones miroir.
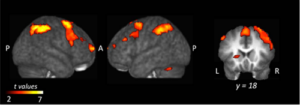
Figure 2. Résultats de l’étude IRMf, publiés dans le journal PlosOne courant 2013, qui permettent de localiser les activations BOLD liées à l’apprentissage social chez l’homme.
Les études sur le système miroir nous informent que les réseaux cérébraux impliqués dans l’expérience des événements (moteurs, sensoriels, émotionnels,…) sont aussi impliqués pendant l’observation d’autres individus en train de vivre la même chose (Wicker et al., 2003 ; Singer et al., 2004 ; Keysers et al., 2004; voir aussi Gallese & Goldman, 1998). Selon ce principe, notre étude en IRMf démontre que, parce que le système fronto-striatal est impliqué dans l’apprentissage individuel (par essai et erreur), il l’est également dans l’apprentissage par observation par le biais d’une résonance motrice. En outre, cette étude nous a permis d’identifier des zones du cerveau spécifiquement activées pendant l’observation des erreurs d’autrui. Dans une étude précédente (cf. Projet ISCI 1, CIR 2012; Monfardini E, Gaveau V, Boussaoud D, Hadj-Bouziane F & Meunier M, 2012. Front. Neurosci. 6:127. doi: 10.3389/ fnins.2012.00127), nous avons démontré que les hommes tirent beaucoup plus de bénéfice des échecs des autres que des leurs propres. Cet effet est stable et présent chez tous les individus testés. Cela pourrait témoigner d’un biais social connu en psychologie sociale et en neuroéconomie : l’homme a tendance à penser que quand les autres font une erreur c’est de leur faute, alors que quand il commet la même erreur, il est simplement victime de la situation (biais « acteur-observateur »). Nos résultats en IRMf identifient les bases cérébrales de ce biais cognitif dans le cortex frontal médial postérieur (pMFC), l’insula antérieure et la partie postérieure du sillon temporal supérieur (pSTS).
5. Indicateurs de R&D
L’article décrivant les résultats de cette étude chez l’homme a été publié courant 2013 dans une revue scientifique internationale (référence : Monfardini E, Gazzola V, Boussaoud D, Brovelli A, Keysers C, Wicker B, 2013. Vicarious Neural Processing of Outcomes during Observational Learning. PLoSONE 8(9): e73879. doi:10.1371/journal.pone.0073879).
6. Acquisition de connaissances
Ainsi, en raison de l’implication du système fronto-striatal dans l’apprentissage individuel (par essai et erreur), que nous avons pu observer lors de cette étude en IRMf, nous pouvons dire qu’il est également impliqué dans l’apprentissage par observation par le biais d’une résonance motrice. Par ailleurs, cette étude nous a permis d’identifier des zones du cerveau spécifiquement activées pendant l’observation des erreurs d’autrui. Dans une étude précédente (cf. Projet ISCI 1, CIR 2012; Monfardini et al., 2012), nous avons démontré que les hommes tirent beaucoup plus de bénéfice des échecs des autres que des leurs propres. Cet effet est stable et présent chez tous les individus testés. Cela pourrait témoigner d’un biais social connu en psychologie sociale et en neuroéconomie : l’homme a tendance à penser que quand les autres font une erreur c’est de leur faute, alors que quand il commet la même erreur, il est simplement victime de la situation (biais « acteur-observateur »). Nos résultats en IRMf identifient les bases cérébrales de ce biais cognitif dans le cortex frontal médial postérieur (pMFC), l’insula antérieure et la partie postérieure du sillon temporal supérieur (pSTS).
7. Ressources humaines
| Nom | Fonction dans le projet | Nbre heures/jours consacrées à la Phase N | Nb total d’heures/jours affectées au projet |
| Elisabetta Monfardini | Chef de projet | Traitement des résultats et écriture de l’article (2014-2015) | 260 heures |
| Jacques Fradin | Directeur de recherche | Traitement des résultats et écriture de l’article (2014-2015) | 30 heures |
8. Références bibliographiques
Asch, S.E. (1956). Studies of independence and conformity: I. A minority of one against a unanimous majority. Psychological Monographs 70(9, Whole No. 416).
Bounoua, L., Cury, F., Monfardini, E., Meunier, M., Boussaoud, D., Brovelli, A. & Huguet, P. (2009). Learning by observation: The role of social comparison and related ability inferences. 16th Annual ESCOP conference, Jagiellonian University, Krakow.
Castro, L. & Toro, M. (2004). The evolution of culture: From primate social learning to human culture. Proceedings of the National Academy of Sciences, 101: 10235-10240.
Dindo, M., Whiten, A. & de Waal, F. B. M. (2009). Social Facilitation of Exploratory Foraging Behavior in Capuchin Monkeys (Cebus Apella). American Journal of Primatology, 71: 419–26.
Ferrari, P.F., Maiolini, C., Addessi, E., Fogassi, L. & Visalberghi, E. (2005). The observation and hearing of eating actions activates motor programs related to eating in macaque monkeys. Behavior Brain Research 161, 95-101.
Fujii, N., Hihara, S. & Iriki A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PLoS ONE 2:e397.
Fujii, N., Hihara, S. & Iriki, S. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience 3, 250-60.
Galef, B.G. & Laland, K.N. (2005). Social learning in animals: empirical studies and theoretical models. Bioscience 55, 489–499.
Galloway, A.T., Addessi, E., Fragaszy, D. & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchin monkeys (Cebus apella): does it involve behavioral coordination? International Journal of Primatology 26, 175–183.
Guerin, B. & Innes, J. (2009). Social facilitation. Cambridge University Press.
Huguet, P., Galvaing, M.P., Monteil, J.M. & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of Personality and Social Psychology 77, 1011-25.
Martin, R. (1998). Majority and minority influence using the afterimage paradigm: A series of attempted replications. Journal of Experimental Social Psychology 34, 1-26.
Monfardini, E., Brovelli, A., Boussaoud, D., Takerkart, S. & Wicker, B. (2008). I learned from what you did: Retrieving visuomotor associations learned by observation. Neuroimage 42, 1207- 13.
Monfardini, E., Gazzola, V., Boussaoud, D., Brovelli, A., Keysers, C. & Wicker, B. (2013). Vicarious Neural Processing of Outcomes during Observational Learning. PLoSONE 8(9): e73879. doi:10.1371/journal.pone.0073879
Visalberghi, E. & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour 60, 69–76.
Visalberghi, E. & Addessi, E. (2001). Acceptance of novel foods in Cebus apella: Do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour 62, 567– 576.
Visalberghi, E. & Fragaszy, D.M. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour 49, 1089-1095.
| NOM DU PROJET : ISCI7 La modulation sociale du taux de cortisol chez le singe. |
| Date de début : 2011 | Date de fin : Prévu 2015 |
| R. Psychologie – Neurosciences |
1. Objectifs du projet
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant les neurosciences cognitives, la psychologie sociale, l’éthologie comparée et la neuroendocrinologie. Plus particulièrement, ces travaux portent sur l’influence des interactions sociales sur le fonctionnement cognitif et cérébral individuel. Cela à travers les méthodes scientifiques des neurosciences fondamentales et de la psychologie (Ochsner & Lieberman, 2001).
Le présent axe de recherche s’inscrit dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres. Dans l’approche proposée, le Positionnement Grégaire aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études décrites ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est phylogénétiquement ancien. Étudier cette influence nous permettra donc d’explorer nos hypothèses en termes de cognition sociale et du fonctionnement cérébral qui en est responsable. D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Le but du présent projet de recherche est d’explorer la modulation sociale de la cognition, les mécanismes qui la gouvernent, son évolution au cours de la phylogénèse, ainsi que ses substrats neuronaux. Il vise en particulier à répondre aux questions suivantes :
– Sommes-nous en quelque sorte ‘programmés’ pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (cf. Fig.1, volet Fondamental) ;
– Quelles modulations sociales sont des tendances phylogénétiquement anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes? (cf. Fig.1, volet Fondamental) ;
– Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale? (cf. Fig.1, volet Bases Neurales) ;
– Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions sont abordées via trois approches complémentaires :
– Une approche comportementale comparée confrontant le comportement de l’homme à celui de l’animal mis dans une situation similaire ; (cf. Projets ISCI2 et ISCI3)
– Deux techniques d’imagerie cérébrale humaine : la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) appliquées au macaque et à l’homme (toutes deux permettent une visualisation non invasive de l’activité du cerveau) ; cf. Projets ISCI4, ISCI6 et ISCI8)
– L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc. (cf. ISCI7).
D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est simplement la position sociale (« social ranking », en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres (« social comparison »)
Dans ce cadre, le présent projet s’intéresse plus particulièrement à la modulation du taux de cortisol (manifestation du stress) chez le singe en situation sociale.
2. État de l’art
La psychologie sociale a fourni des données expérimentales sur l’influence de la socialité sur le fonctionnement cognitif individuel. Ces études témoignent du fait que la simple présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives, consistant soit à les améliorer, soit à les détériorer (pour une revue récente de la littérature, voir Guerin & Innes, 2009). Le phénomène de modulation sociale concerne une large palette de capacités cognitives : des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements), aux plus complexes comme les capacités attentionnelles (e.g. dans la tâche de Stroop, Huguet et al., 1999), la prise de décision (Asch, 1956 ; Martin, 1998 ; ), ou l’apprentissage social de règles (Bounoua et al., 2009). En résumé, des situations sociales relativement simples peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales ou cognitives.
Les effets d’influence sociale ne sont pas une prérogative humaine. Des phénomènes de facilitation sociale ont aussi été démontrés chez les primates non-humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire. Par exemple, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Au niveau cérébral, une seule série d’études a jusqu’à présent donné quelques éléments pour répondre à cette question. Les auteurs (Fujii et al., 2007 ; 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone.
Comme nous l’avons précédemment dit, l’apprentissage social est également une composante fondamentale de la cognition humaine. Apprendre par observation d’autrui est vital pour l’homme (Castro & Toro, 2004). Sans cet apprentissage, chaque enfant devrait réapprendre chaque comportement laborieusement par lui-même et aucune transmission de savoir-faire ne serait possible. Selon Bandura (1965), ce type d’apprentissage est caractérisé par quatre étapes: l’attention, le maintien en mémoire, la reproduction du geste, et la motivation. La première étape, l’attention, est un processus crucial et peut être fortement influencé par les caractéristiques du modèle. Il a été démontré que des caractéristiques telles que l’attractivité, la similitude et la compétence perçue améliorent l’efficacité de l’apprentissage par observation (Brewer & Wann, 1998). En particulier, la similitude entre acteur et observateur serait la clé d’une transmission sociale des connaissances efficace. Adultes comme enfants savent en effet bénéficier de l’observation d’individus dont les performances sont légèrement supérieures quand ils s’agit d’apprendre des associations stimulus-récompense, mais au contraire ne bénéficient pas de l’observation d’individus perçus comme trop experts (Bounoua et al., 2009). Des récentes données de neurosciences corroborent ces données, en montrant une modulation des mécanismes cérébraux responsables de l’apprentissage social selon le dégrée de similarité et/ou affinité perçu entre modèle et apprenti (Mobbs et al., 2009 ; Leng & Zhou, 2010).
Pour le présent projet, nous avons souhaité étudier les phénomènes physiologiques liées aux interactions sociales telles que les rapports affiliatifs, les rapports de dominance et leur interface avec le fonctionnement cognitif de l’individu. L’étude de ces phénomènes, et notamment de l’apparition du stress physiologique dans les interactions sociales, est en effet particulièrement importante dans le cadre de la compréhension des mécanismes neuropsychologiques en jeu dans les situations d’apprentissage social.
3. Aléas, incertitudes scientifiques, verrous technologiques
L’étude neuroscientifique de la modulation sociale de la cognition et, plus en général, de la sphère sociale de notre existence, présente un grand intérêt mais aussi des défis techniques considérables. Premier élément, le contrôle d’un grand nombre de variables qui interviennent dans nos rapports sociaux et qui influencent notre « positionnement grégaire ». La complexité de ce domaine de recherche implique la multiplication des variables expérimentales à inclure dans nos protocoles expérimentaux et, par conséquent, des coûts considérables en termes de temps et de prix d’examens (notamment pour ce qui concerne les études neuroscientifiques d’imagerie). Cependant, les collaborations scientifiques sur lesquelles ces études reposent, ainsi que les publications scientifiques nationales et internationales qui en découlent, nous permettent de faire face aux problèmes liés à l’originalité de ce domaine d’étude. En effet, les résultats de ces études ont déjà fait l’objet de plusieurs publications dans des journaux scientifiques internationaux (Plos One, NeuroImage, Frontiers In Neurosciences, etc.) et positionnent le Pôle de Recherche de l’IME dans le panorama scientifique international aussi bien dans le domaine de la Psychologie Cognitive que des Neurosciences.
L’étude ISCI7 a impliqué l’utilisation d’un modèle animal, le primate non-humain. Ceci rentre dans le cadre d’une approche comparative finalisée à isoler les composantes fondamentales des comportements humains, qui s’expriment indépendamment de la culture et du langage. En particulier, le choix du modèle primate est motivé par l’intérêt que nous portons aux hiérarchies sociales et au positionnement grégaire. Or, les singes macaques passent la majeure partie de leur vie éveillée en présence de leurs congénères, et les rapports qu’ils instaurent au sein d’un groupe sont gérés par de fortes hiérarchies sociales. L’utilisation d’un modèle animal comporte des coûts et des aléas tout à fait différents de ceux auxquels on fait face pour les études chez l’homme. Tout d’abord, l’hébergement des animaux requiert du personnel et des infrastructures très coûteuses. Ensuite, la réalisation de protocoles expérimentaux impliquant des tâches cognitives requiert l’entrainement des animaux, qui se fait de façon progressive souvent sur des longues périodes (1 à 6 mois). Cet entrainement, qui comporte des travers les méthodes de conditionnement opérant bien connues en éthologie et psychologie comparative, demande des compétences particulières de la part du personnel impliqué dans l’étude. Le projet ISCI7 a été mené avec succès en collaboration avec le Centre de Recherche en Neurosciences de Lyon, qui possède une animalerie primate complètement rénovée et pouvant héberger un nombre important d’animaux. Le personnel INSERM qui y travaille a assuré le soin des animaux et leur hébergement conformément aux directives européennes pendant tout le déroulement du projet.
4. Travaux R&D réalisés, démarche expérimentale
La « Neuroendocrinologie Sociale » est un domaine émergent de recherche interdisciplinaire qui combine les théories et les approches de la neuroendocrinologie comportementales et la psychologie sociale et de la personnalité. Les chercheurs dans ce domaine dédient leurs activités à la compréhension des interactions entre les hormones, l’environnement et le comportement social. Nous avons donc commencé à explorer ce champ d’études qui donne plusieurs opportunités d’étude des phénomènes physiologiques liées aux interactions sociales telles que les rapports affiliatifs, les rapports de dominance et leur interface avec le fonctionnement cognitif de l’individu.
Plus particulièrement, nous nous attachons actuellement à tester un protocole d’étude des concentrations plasmatiques de cortisol (CORT) et d’hormone corticotrophine (ACTH), qui sont des indicateurs de l’activité hormonale de l’axe HPA (hypothalamus-hypophyse-corticosurrénale) et donc fortement liés à des manifestations de stress (Nelson, 1995), au cours des situations de facilitation/inhibition sociale. Le but de l’étude est d’explorer l’hypothèse que la concentration plasmatique de cortisol puisse varier en fonction de la situation sociale vécue, qui a son tour influence les performances cognitives individuelles. Cinq singes macaques rhésus sont nos sujets expérimentaux. Le protocole consiste à prélever des échantillons de sang (4ml de sang de la veine saphène) avant (PRE) et après (POST) la situation expérimentale, qui consiste à exécuter une tâche cognitive d’apprentissage dans trois situations sociales différentes : isolation sociale (Alone), présence d’un congénère familier (Fam), présence d’un congénère non familier (Nfam, donc de rang social inconnu; cf. Figure 2). Les échantillons, qui sont analysés par un cabinet d’analyses médicales, nous ont permis d’obtenir des données préliminaires. Ces dernières nous donnent des indications intéressantes (1) sur la grande variabilité inter-individuelle du taux de cortisol prélevé avant la situation expérimentale (PRE), qui peut être corrélée au rang social de chacun des animaux (Martin & Wrangham, 2004) ; (2) sur le stress engendré par les situations sociales décrites ci-dessus.
| Subj | Avge PRE | POST Fam | POST Alone | POST Nfam |
| CANELLE | 26,2 | 24,2 | 27,3 | 29,6 |
| CASSIS | 26,2 | 33,9 | 27,3 | 34,0 |
| CELIA | 21,9 | 32,1 | 37,0 | 40,0 |
| CEYRESTE | 9,9 | 20,1 | 27,0 | 21,4 |
| CIOTAT | 23,8 | 32,2 | 36,8 | 41,0 |
| mean | 21,6 | 28,5 | 31,1 | 33,2 |
| Sem | 3,0 | 2,7 | 2,4 | 3,6 |
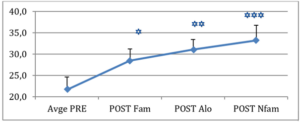
Figure 2. Résultats de l’étude ISCI7, permettant d’explorer le niveau de cortisol plasmatique associé à des situations sociales de nature différente (Fam = présence d’un congénère familier; Nfam = présence d’un congénère inconnu ; Alo = isolation sociale).
5. Indicateurs de R&D
Les résultats de l’étude ISCI7 seront intégrés dans l’article de l’étude ISCI4. Leur publication est le prochain objectif pour 2014.
6. Acquisition de connaissances
Les résultats de cette étude semblent montrer que le niveau de stress ne change pas en fonction de la situation sociale testée. Or, en présence d’autrui, les performances sont améliorées par rapport à quand les animaux sont testés seuls. Ces résultats sont complémentaires à ceux des études ISCI3 et ISCI4, qui démontrent que (1) en présence d’autrui, les performances sont améliorées aussi bien dans le cas d’une tâche maitrisée que dans une tâche non maîtrisée, et aussi bien quand l’autre individu présent travaille que quand il est passif; et (2) que la simple présence d’autrui lors de la réalisation d’une tâche serait capable de moduler l’activité cérébrale au sein du réseau fronto-pariétal de l’attention. Les autres changeraient notre capacité à prêter attention à une tâche, et ceci en agissant sur nos mécanismes cérébraux « de base »). ISCI7 apporte une pierre à l’édifice de la compréhension du fonctionnement du cerveau en interaction sociale.
Les résultats de l’étude ISCI7 seront intégrés dans l’article de l’étude ISCI4, en révision dans le journal scientifique international Cerebral Cortex. (Monfardini E, Redouté J, Hadj-Bouziane F, Hynaux C, Fradin J, Huguet P, Costes N, Meunier M (under revision). Others’ sheer presence boosts brain activity in the attention (but not the motivation) network.
7. Ressources humaines
| Nom | Fonction dans le projet | Nbre heures/jours consacrées à la Phase N | Nb total d’heures/jours affectées au projet |
|
Elisabetta Monfardini
|
Chef de projet | Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | 260 heures |
| Jacques Fradin | Directeur de recherche | Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | 29 heures |
8. Références bibliographiques
Asch, S.E. (1956). Studies of independence and conformity: I. A minority of one against a unanimous majority. Psychological Monographs 70(9, Whole No. 416).
Bandura, A. (1965). Influence of models’ reinforcement contingencies on the acquisition of imitative responses. Journal of Personnality and Social Psychology, 1, 589-595.
Bounoua, L., Cury, F., Monfardini, E., Meunier, M., Boussaoud, D., Brovelli, A. & Huguet, P. (2009). Learning by observation: The role of social comparison and related ability inferences. 16th Annual ESCOP conference, Jagiellonian University, Krakow.
Dindo, M., Whiten, A. & de Waal, F. B. M. (2009). Social Facilitation of Exploratory Foraging Behavior in Capuchin Monkeys (Cebus Apella). American Journal of Primatology, 71, 419–26.
Ferrari, P.F., Maiolini, C., Addessi, E., Fogassi, L. & Visalberghi, E. (2005). The observation and hearing of eating actions activates motor programs related to eating in macaque monkeys. Behavior Brain Research 161, 95-101.
Fujii, N., Hihara, S. & Iriki A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PLoS ONE 2:e397.
Fujii, N., Hihara, S. & Iriki, S. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience 3, 250-60.
Galloway, A.T., Addessi, E., Fragaszy, D. & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchin monkeys (Cebus apella): does it involve behavioral coordination? International Journal of Primatology 26, 175–183.
Guerin, B. & Innes, J. (2009). Social facilitation. Cambridge University Press.
Huguet, P., Galvaing, M.P., Monteil, J.M. & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of Personality and Social Psychology 77, 1011-25.
Martin, N.M. & Richard, W.W. (2004). Dominance, cortisol and stress in wild chimpanzees (Pan troglodytes schweinfurthii). Behavioral Ecology and Sociobiology, 55, 332–340.
Martin, R. (1998). Majority and minority influence using the afterimage paradigm: A series of attempted replications. Journal of Experimental Social Psychology 34, 1-26.
Nelson, R.J. (1995). An introduction to behavioral endocrinology. Sinauer, Sunderland, Mass.
Visalberghi, E. & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour 60: 69–76.
Visalberghi, E. & Addessi, E. (2001). Acceptance of novel foods in Cebus apella: Do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour 62, 567– 576.
Visalberghi, E. & Fragaszy, D.M. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour 49, 1089-1095.
| NOM DU PROJET : ISCI8- Les bases neurales du positionnement grégaire chez le primate humain et non-humain: Mise au point méthodologique pour l’étude de l’activité cérébrale au repos. |
| Date de début : 2013 | Date de fin : 2014 |
| R. Psychologie – Neurosciences |
1. Objectifs du projet
L’ensemble des recherches « ISCI » s’inscrit dans le cadre d’un projet pluridisciplinaire reliant les neurosciences cognitives, la psychologie sociale, l’éthologie comparée et la neuroendocrinologie. Plus particulièrement, ces travaux portent sur l’influence des interactions sociales sur le fonctionnement cognitif et cérébral individuel. Cela à travers les méthodes scientifiques des neurosciences fondamentales et de la psychologie (Ochsner & Lieberman, 2001).
Le présent axe de recherche s’inscrit dans l’intérêt que l’IME porte au « Positionnement Grégaire » (PG), décrit comme la position sociale (en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) qu’un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres. Dans l’approche proposée, le Positionnement Grégaire aurait un impact sur la genèse de certaines psychopathologies comme le trouble obsessionnel compulsif ou la dépression. Les études décrites ci-dessous démontrent que les relations dans un groupe social exercent une influence drastique (positive ou négative) sur les performances cognitives et comportementales des individus qui le composent, et que ce phénomène est phylogénétiquement ancien. Étudier cette influence nous permettra donc d’explorer nos hypothèses en termes de cognition sociale et du fonctionnement cérébral qui en est responsable. D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale. L’intégration des contenus issus de présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Le but du présent projet de recherche est d’explorer la modulation sociale de la cognition, les mécanismes qui la gouvernent, son évolution au cours de la phylogénèse, ainsi que ses substrats neuronaux. Il vise en particulier à répondre aux questions suivantes :
– Sommes-nous en quelque sorte ‘programmés’ pour modifier nos comportements, décisions et pensées en fonction de la présence des autres et des interactions que nous instaurons avec eux? (cf. Fig.1, volet Fondamental) ;
– Quelles modulations sociales sont des tendances phylogénétiquement anciennes que l’homme, malgré son langage et sa sophistication culturelle et intellectuelle, partage avec d’autres espèces comme les singes? (cf. Fig.1, volet Fondamental) ;
– Quels sont les circuits cérébraux et la dynamique neuronale sous-jacents à ces phénomènes d’influence sociale? (cf. Fig.1, volet Bases Neurales) ;
– Quelles implications l’influence sociale a sur la genèse de certaines psychopathologies comme la dépression ou le trouble obsessionnel compulsif, la dépression, le trouble d’anxiété généralisé, l’anxiété sociale, les troubles de la personnalité antisociale ou paranoïaque ? (cf. Fig.1, volet Clinique).
Ces questions sont abordées via trois approches complémentaires :
– Une approche comportementale comparée confrontant le comportement de l’homme à celui de l’animal mis dans une situation similaire ; (cf. Projets ISCI2 et ISCI3)
– Deux techniques d’imagerie cérébrale humaine : la tomographie par émission de positrons (TEP) et l’imagerie par résonnance magnétique (IRMf) appliquées au macaque et à l’homme (toutes deux permettent une visualisation non invasive de l’activité du cerveau) ; cf. Projets ISCI4, ISCI6 et ISCI8)
– L’approche expérimentale du domaine émergent de la ‘neuroendocrinologie sociale’, qui cherche à comprendre les liens entre la cognition sociale et les principales hormones produites par le système nerveux : cortisol, testostérone, ocytocine, etc. (cf. ISCI7).
D’un point de vue applicatif, les résultats des recherches ISCI seront, d’une part, intégrés dans les cycles de formations déjà proposés par l’IME et, d’autre part, feront l’objet de formations spécifiques visant à améliorer les capacités cognitives des individus en contexte professionnel ou scolaire à l’aide des connaissances neuroscientifiques sur le fonctionnement du cerveau en interaction sociale.
L’intégration des contenus issus des présentes recherches et des besoins formatifs des populations visées par l’IME (managers, professionnels de la santé, particuliers, etc.) fait l’objet d’un projet de laboratoire commun entre le CNRL et l’IME dénommé NeuroTeach (demande de subvention de l’ANR en cours), et qui est destiné à valoriser les produits de la recherche des deux organisations dans le cadre d’applications thérapeutiques, pédagogiques, managériales et industrielles.
Figure 1. Organisation du volet de recherche « Positionnement Grégaire » (PG) développé au sein de l’IME. Le PG est simplement la position sociale (« social ranking », en terme de dominance/soumission et de centralité/marginalité d’un membre dans son groupe) que un individu occupe dans son milieu et la comparaison sociale qu’il établi avec les autres (« social comparison »)
L’objectif du présent projet est d’étudier la connectivité fonctionnelle intrinsèque d’individus ayant un Positionnement Grégaire (i.e. position sociale, en terme de dominance-soumission et centralité-marginalité) diffèrent, à l’aide de l’imagerie par Résonance Magnétique fonctionnelle et au repos (rs-IMRf). Une telle étude permettrait de mieux cerner les mécanismes en jeu dans les interactions sociales, notamment de type dominance-soumission.
2. État de l’art
La psychologie sociale a fourni des données expérimentales sur l’influence de la socialité sur le fonctionnement cognitif individuel. Ces études témoignent du fait que la simple présence (active ou passive) d’autrui lors de l’accomplissement d’une tâche a des conséquences directes sur nos performances cognitives, consistant soit à les améliorer, soit à les détériorer (pour une revue récente de la littérature, voir Guerin & Innes, 2009). Le phénomène de modulation sociale concerne une large palette de capacités cognitives : des plus simples comme les performances motrices (e.g. la vitesse à laquelle on effectue une séquence de mouvements), aux plus complexes comme les capacités attentionnelles (e.g. dans la tâche de Stroop, Huguet et al., 1999), la prise de décision (Asch, 1956 ; Martin, 1998 ; ), ou l’apprentissage social de règles (Bounoua et al., 2009). En résumé, des situations sociales relativement simples peuvent avoir des effets impressionnants sur une large gamme de réponses comportementales ou cognitives.
Les effets d’influence sociale ne sont pas une prérogative humaine. Des phénomènes de facilitation sociale ont aussi été démontrés chez les primates non-humains. Un des comportements particulièrement sensible à l’influence sociale est la prise alimentaire. Par exemple, les singes capucins qui regardent les autres membres du groupe manger de la nourriture augmentent leur propre prise de nourriture (Dindo et al., 2009 ; Galloway et al., 2005 ; Visalberghi & Addessi 2000, 2001; Visalberghi & Fragaszy, 1995). Cette facilitation s’exerce non seulement à la vue des autres membres qui se nourrissent, mais aussi à la seule écoute des bruits qu’ils génèrent (Ferrari et al., 2005).
Au niveau cérébral, une seule série d’études a jusqu’à présent donné quelques éléments pour répondre à cette question. Les auteurs (Fujii et al., 2007 ; 2008) ont enregistré l’activité neuronale du cortex pariétal de deux singes partageant un espace social. Quand les singes étaient assis l’un près de l’autre, mais sans interagir, l’activité du cortex pariétal de chacun reflétait uniquement les mouvements propres à l’animal. En revanche, en situation d’interaction sociale (par exemple, quand un seul morceau de nourriture était placé sur la table et que les deux singes pouvaient l’atteindre), l’activité du cortex pariétal reflétaient non seulement les mouvements propres à l’animal, mais aussi les mouvements de l’autre. Cette étude pionnière démontrait pour la première fois que les facteurs sociaux affectent l’activité du cerveau, et cette modulation est détectable jusqu’au niveau le plus unitaire, celui du neurone.
Comme nous l’avons précédemment dit, l’apprentissage social est également une composante fondamentale de la cognition humaine. Apprendre par observation d’autrui est vital pour l’homme (Castro & Toro, 2004). Sans cet apprentissage, chaque enfant devrait réapprendre chaque comportement laborieusement par lui-même et aucune transmission de savoir-faire ne serait possible. Selon Bandura (1965), ce type d’apprentissage est caractérisé par quatre étapes: l’attention, le maintien en mémoire, la reproduction du geste, et la motivation. La première étape, l’attention, est un processus crucial et peut être fortement influencé par les caractéristiques du modèle. Il a été démontré que des caractéristiques telles que l’attractivité, la similitude et la compétence perçue améliorent l’efficacité de l’apprentissage par observation (Brewer & Wann, 1998). En particulier, la similitude entre acteur et observateur serait la clé d’une transmission sociale des connaissances efficace. Adultes comme enfants savent en effet bénéficier de l’observation d’individus dont les performances sont légèrement supérieures quand ils s’agit d’apprendre des associations stimulus-récompense, mais au contraire ne bénéficient pas de l’observation d’individus perçus comme trop experts (Bounoua et al., 2009). Des récentes données de neurosciences corroborent ces données, en montrant une modulation des mécanismes cérébraux responsables de l’apprentissage social selon le dégrée de similarité et/ou affinité perçu entre modèle et apprenti (Mobbs et al., 2009 ; Leng & Zhou, 2010).
Par ailleurs, la neuropsychologie s’intéresse depuis quelques années au fait que les personnes pensent ou se comportent différemment l’une de l’autre soit ancré dans les grandes différences interindividuelles que l’on retrouve dans leur cerveau (e.g. Frost et Goebel, 2012). Etudier les connections qui existent entre les différentes aires d’un cerveau (i.e. le connectome) semble être parmi les meilleures façons de dénicher les caractéristiques spécifiques à chaque cerveau (Seung, 2012).
Les données d’imagerie par Résonance Magnétique Fonctionnelle obtenues dans des situations de repos (rs-IRMf) offrent la possibilité d’évaluer cette connectivité, qui est souvent appelée connectivité fonctionnelle intrinsèque (Fox et Raichle, 2007). Il a été démontré que des différences interindividuelles dans la connectivité fonctionnelle intrinsèque, par exemple, peuvent prédire la grande variabilité que l’on retrouve au niveau des performances cognitives chez l’homme (Andrews-Hanna et al, 2007; Seeley et al, 2007 ; van den Heuvel et al, 2009), ainsi que la sévérité des symptômes de troubles neuropsychiatriques (Fox et Greicius, 2010 ; Greicius, 2008). Ceci suggère que des nouvelles connaissances sur les mécanismes neuronaux sous-jacents à la variabilité individuelle des fonctions humaines pourraient découler d’une meilleure compréhension de la distribution spatiale de la connectivité intrinsèque dans le cerveau.
L’activité cérébrale au repos, plus connue sous le nom de ‘resting state network’, a fait l’objet de nombreuses investigations en imagerie cérébrale au cours de cette dernière décennie. Les travaux réalisés en IRMf au repos (rs-IRMf) démontrent que, d’un point de vue physiologique, ces réseaux sont caractérisés par la présence de fluctuations basse fréquence du signal allant de 0.01 à 0.1 Hz et principalement, mais pas exclusivement, détectables au niveau du cortex cérébral.
L’objectif de ce projet est donc d’étudier la connectivité fonctionnelle intrinsèque d’individus ayant un Positionnement Grégaire (i.e. position sociale, en terme de dominance-soumission et centralité-marginalité) diffèrent, à l’aide de l’imagerie par Résonance Magnétique fonctionnelle au repos (rs-IMRf). Notre hypothèse est que le cerveau d’individus caractérisés par une forte dominance ou soumission présenterait des différences en terme de ‘connections’ fonctionnelles entre les aires cérébrales par rapports aux individus assertifs. Ces différences, selon notre hypothèse, seraient surtout localisés au niveau du réseau cérébral dit ‘limbique’ (Mueller et al, 2013) et de ses connections avec les aires corticales frontales.
3. Aléas, incertitudes scientifiques, verrous technologiques
L’étude neuroscientifique de la modulation sociale de la cognition et, plus en général, de la sphère sociale de notre existence, présente un grand intérêt mais aussi des défis techniques considérables. Premier élément, le contrôle d’un grand nombre de variables qui interviennent dans nos rapports sociaux et qui influencent notre « positionnement grégaire ». La complexité de ce domaine de recherche implique la multiplication des variables expérimentales à inclure dans nos protocoles expérimentaux et, par conséquent, des coûts considérables en termes de temps et de prix d’examens (notamment pour ce qui concerne les études neuroscientifiques d’imagerie). Cependant, les collaborations scientifiques sur lesquelles ces études reposent, ainsi que les publications scientifiques nationales et internationales qui en découlent, nous permettent de faire face aux problèmes liés à l’originalité de ce domaine d’étude. En effet, les résultats de ces études ont déjà fait l’objet de plusieurs publications dans des journaux scientifiques internationaux (Plos One, NeuroImage, Frontiers In Neurosciences, etc.) et positionnent le Pôle de Recherche de l’IME dans le panorama scientifique international aussi bien dans le domaine de la Psychologie Cognitive que des Neurosciences.
Pour l’étude qui nous intéresse, un des verrous technologiques qui se présente à nous est le fait que le rs-IRMf constitue un enjeu méthodologique majeur et requiert un ensemble de méthodes d’analyses sophistiquées. L’année 2013 nous a permis de peaufiner ces méthodes et de mettre au point un système permettant d’acquérir des données d’IRMf chez le singe macaque vigile en situation de repos.
L’étude ISCI8 a impliqué l’utilisation d’un modèle animal, le primate non-humain. Ceci rentre dans le cadre d’une approche comparative finalisée à isoler les composantes fondamentales des comportements humains, qui s’expriment indépendamment de la culture et du langage. D’un point de vue neuroscientifique, comparer le cerveau de différentes espèces nous permet d’évaluer la phylogenèse de la neuroanatomie, et ouvre la voie à des méthodes
plus invasives d’étude. En particulier, le choix du modèle primate est motivé par l’intérêt que nous portons aux hiérarchies sociales et au positionnement grégaire. Or, les singes macaques passent la majeure partie de leur vie éveillée en présence de leurs congénères, et les rapports qu’ils instaurent au sein d’un groupe sont gérés par de fortes hiérarchies sociales. L’utilisation d’un modèle animal comporte des coûts et des aléas tout à fait différents de ceux auxquels on fait face pour les études chez l’homme. Tout d’abord, l’hébergement des animaux requiert du personnel et des infrastructures très coûteuses. Ensuite, la réalisation de protocoles expérimentaux impliquant des tâches cognitives requiert l’entrainement des animaux, qui se fait de façon progressive souvent sur des longues périodes (1 à 6 mois). Cet entrainement, qui comporte des travers les méthodes de conditionnement opérant bien connues en éthologie et psychologie comparative, demande des compétences particulières de la part du personnel impliqué dans l’étude. Le projet ISCI8 a été mené avec succès en collaboration avec le Centre de Recherche en Neurosciences de Lyon, qui possède une animalerie primate complètement rénovée et pouvant héberger un nombre important d’animaux. Le personnel INSERM qui y travaille a assuré le soin des animaux et leur hébergement conformément aux directives européennes pendant tout le déroulement du projet.
4. Travaux R&D réalisés, démarche expérimentale
Le fait que les personnes pensent ou se comportent différemment l’une de l’autre est ancrée dans les grandes différences interindividuelles que l’on retrouve dans leur cerveau (e.g. Frost et Goebel, 2012). Etudier les connections qui existent entre les différentes aires d’un cerveau (i.e. le connectome) semble être parmi les meilleures façons de dénicher les caractéristiques spécifiques à chaque cerveau (Seung, 2012).
Les données d’imagerie par Résonance Magnétique Fonctionnelle obtenues dans des situations de repos (rs-IRMf) offrent la possibilité d’évaluer cette connectivité, qui est souvent appelée connectivité fonctionnelle intrinsèque (Fox et Raichle, 2007). Il a été démontré que des différences interindividuelles dans la connectivité fonctionnelle intrinsèque, par exemple, peuvent prédire la grande variabilité que l’on retrouve au niveau des performances cognitives chez l’homme (Andrews-Hanna et al, 2007; Seeley et al, 2007; van den Heuvel et al, 2009), ainsi que la sévérité des symptômes de troubles neuropsychiatriques (Fox et Greicius, 2010; Greicius, 2008). Ceci suggère que des nouvelles connaissances sur les mécanismes neuronaux subjacents à la variabilité individuelle des fonctions humaines pourraient découler d’une meilleure compréhension de la distribution spatiale de la connectivité intrinsèque dans le cerveau (figure 2).
L’activité cérébrale au repos, plus connue sous le nom de ‘resting state network’, a fait l’objet de nombreuses investigations en imagerie cérébrale au cours de cette dernière décennie. Les travaux réalisés en IRMf au repos (rs-IRMf) démontrent que, d’un point de vue physiologique, ces réseaux sont caractérisés par la présence de fluctuations basse fréquence du signal allant de 0.01 à 0.1 Hz et principalement, mais pas exclusivement, détectables au niveau du cortex cérébral.
L’objectif de ce projet est d’étudier la connectivité fonctionnelle intrinsèque d’individus ayant un Positionnement Grégaire (i.e. position sociale, en terme de dominance-soumission et centralité-marginalité) diffèrent, à l’aide de l’imagerie par Résonance Magnétique fonctionnelle au repos (rs-IMRf). Notre hypothèse est que le cerveau d’individus caractérisés par une forte dominance ou soumission présenterait des différences en terme de ‘connections’ fonctionnelles entre les aires cérébrales par rapports aux individus assertifs. Ces différences, selon notre hypothèse, seraient surtout localisés au niveau du réseau cérébral dit ‘limbique’ (Mueller et al, 2013) et de ses connections avec les aires corticales frontales. Nos hypothèses seront testées à la fois chez l’homme et le primate non-humain, afin d’isoler les mécanismes neurobiologiques fondamentaux capables de prédire le Positionnement Grégaire d’un individu.
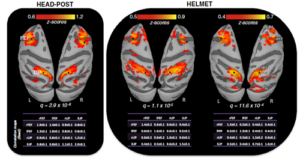
Figure 2. Résultats de l’étude méthodologique de rs-IRMf chez 3 singes macaques (à paraître dans le journal NeuroImage).
5. Indicateurs de R&D
Un premier article décrivant notre méthodologie a été publié dans le journal scientifique international « NeuroImage ». (Hadj-Bouziane F*, Monfardini E*, Guedj C*, Gardechaux G , Hynaux C , Farnè A, Meunier M (2014). Validating the helmet head restraint method for resting state fMRI in awake monkeys. NeuroImage. 1, 536-43.*co-1st authors).
6. Acquisition de connaissances
Le rs-IRMf est un enjeu méthodologique majeur et requiert un ensemble des méthodes d’analyses sophistiquées. L’année 2013 nous a permis de peaufiner ces méthodes et de mettre au point un système permettant d’acquérir des données d’IRMf chez le singe macaque vigile en situation de repos. Un premier article décrivant notre méthodologie a été publié dans le journal scientifique international « NeuroImage », une reference dans le domaine de la neuroimagerie. Très peu de groupes de recherche dans le monde ont pu mettre en place avec succès l’imagerie chez le primate non-humain, à cause de nombreux difficultés techniques que cela comporte. La publication de notre article nous permet (1) d’être reconnus dans ce domaine d’étude, et ceci à niveau international ; (2) d’assoir une technique qui nous permettra d’étudier les bases neuronales du positionnement grégaire d’une façon précise et rigoureuse et chez une espèce, le singe macaque, qui présente un modèle de socialité fortement basé sur les hiérarchies sociales.
7. Ressources humaines
| Nom | Fonction dans le projet | Tâche relative au projet | Nb total d’heures/jours affectées au projet |
|
Elisabetta Monfardini
|
Chef de projet | Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | 346 heures |
| Jacques Fradin | Mise en œuvre du protocole et traitement des résultats préliminaires (2012-2013) | 29 heures |
8. Références bibliographiques
Andrews-Hanna, J.R., Snyder, A.Z., Vincent, J.L., Lustig, C., Head, D., Raichle, M.E. & Buckner, R.L. (2007). Disruption of large-scale brain systems in advanced aging. Neuron 56, 924–935.
Asch, S.E. (1956). Studies of independence and conformity: I. A minority of one against a unanimous majority. Psychological Monographs 70(9, Whole No. 416).
Bandura, A. (1965). Influence of models’ reinforcement contingencies on the acquisition of imitative responses. Journal of Personnality and Social Psychology, 1, 589-595.
Bounoua, L., Cury, F., Monfardini, E., Meunier, M., Boussaoud, D., Brovelli, A. & Huguet, P. (2009). Learning by observation: The role of social comparison and related ability inferences. 16th Annual ESCOP conference, Jagiellonian University, Krakow.
Dindo, M., Whiten, A. & de Waal, F. B. M. (2009). Social Facilitation of Exploratory Foraging Behavior in Capuchin Monkeys (Cebus Apella). American Journal of Primatology, 71: 419–26.
Ferrari, P.F., Maiolini, C., Addessi, E., Fogassi, L. & Visalberghi, E. (2005). The observation and hearing of eating actions activates motor programs related to eating in macaque monkeys. Behavior Brain Research 161: 95-101.
Fox, M.D. & Greicius, M. (2010). Clinical applications of resting state functional connectivity. Frontiers in Systems Neurosciences, 4, 19.
Fox, M.D. & Raichle, M.E. (2007). Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nature Review Neurosciences 8, 700–711.
Frost, M.A. & Goebel, R. (2012). Measuring structural-functional correspondence: spatial variability of specialised brain regions after macro-anatomical alignment. Neuroimage 59, 1369–1381.
Fujii, N., Hihara, S. & Iriki A. (2007). Dynamic social adaptation of motion-related neurons in primate parietal cortex. PLoS ONE 2:e397.
Fujii, N., Hihara, S. & Iriki, S. (2008). Social cognition in premotor and parietal cortex. Social Neuroscience 3:250-60.
Galloway, A.T., Addessi, E., Fragaszy, D. & Visalberghi, E. (2005). Social facilitation of eating familiar food in tufted capuchin monkeys (Cebus apella): does it involve behavioral coordination? International Journal of Primatology 26: 175–183.
Greicius, M.D. (2007). Dissociable intrinsic connectivity networks for salience processing and executive control. Journal of Neurosciences, 27, 2349–2356.
Greicius, M. (2008). Resting-state functional connectivity in neuropsychiatric disorders. Current Opinion in Neurology, 21, 424–430.
Guedj, C., Hadj-Bouziane, F., Monfardini, E., Gardechaux, G., Hynaux, C., Farné, A. & Meunier, M. (2013). The helmet head restraint system: a viable solution for resting state fMRI in awake monkeys. Poster. NC3Rs Primate Welfare Meeting, London – UK.
Guedj, C., Monfardini, E., Farnè, A., Meunier, M. & Hadj-Bouziane F (2013). Resting state networks in the awake monkey. Poster. Colloque de la Société Française de Neurosciences, Lyon- France.
Guerin, B. & Innes, J. (2009). Social facilitation. Cambridge University Press.
Hadj-Bouziane, F.*, Monfardini, E.*, Guedj, C.*, Gardechaux, G., Hynaux, C., Farnè, A. & Meunier, M. (2014). Validating the helmet head restraint method for resting state fMRI in awake monkeys. NeuroImage, 1, 536-43.*co-1st authors
Huguet, P., Galvaing, M.P., Monteil, J.M. & Dumas, F. (1999). Social presence effects in the Stroop task: further evidence for an attentional view of social facilitation. Journal of Personality and Social Psychology 77: 1011-25.
Martin, R. (1998). Majority and minority influence using the afterimage paradigm: A series of attempted replications. Journal of Experimental Social Psychology 34: 1-26.
Monfardini, E. (2014). Non-invasive neuroimaging in the macaque monkey. EFOR meeting, Paris- France.
Mueller, S., Wang, D., Fox, M.D., Yeo, B.T., Sepulcre, J., Sabuncu, M.R., Shafee, R., Lu, J. & Liu, H. (2013). Individual variability in functional connectivity architecture of the human brain. Neuron, 6;77(3):586-95. doi: 10.1016/j.neuron.2012.12.028.
van den Heuvel, M.P., Stam, C.J., Kahn, R.S. & Hulshoff Pol, H.E. (2009). Efficiency of functional brain networks and intellectual performance. Journal of Neurosciences,. 29, 7619–7624.
Seeley, W.W., Menon, V., Schatzberg, A.F., Keller, J., Glover, G.H., Kenna, H., Reiss, A.L., & Greicius, M. D. (2007). Dissociable Intrinsic Connectivity Networks for Salience Processive and Executive Control. The Journal of Neuroscience, 27(9), 2349-2356; doi: 10.1523/JNEUROSCI.5587-06.2007
Visalberghi, E. & Addessi, E. (2000). Seeing group members eating a familiar food enhances the acceptance of novel foods in capuchin monkeys. Animal Behaviour 60: 69–76.
Visalberghi, E. & Addessi, E. (2001). Acceptance of novel foods in Cebus apella: Do specific social facilitation and visual stimulus enhancement play a role? Animal Behaviour 62 : 567– 576.
Visalberghi, E. & Fragaszy, D.M. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: the role of social context. Animal Behaviour 49: 1089-1095.
